Impact Factor ISSN: 1449-2288

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2009; 5(4):319-330. doi:10.7150/ijbs.5.319 This issue Cite
Research Paper
A male-specific odorant receptor conserved through the evolution of sex pheromones in Ostrinia moth species
Nami Miura1, Tatsuro Nakagawa2, Sadahiro Tatsuki1, Kazushige Touhara2, Yukio Ishikawa1 ![]()
1. Department of Agricultural and Environmental Biology, Graduate School of Agricultural and Life Sciences, University of Tokyo, Tokyo 113-8657, Japan
2. Department of Integrated Biosciences, Graduate School of Frontier Sciences, University of Tokyo, Chiba 277-8562, Japan
Received 2009-3-25; Accepted 2009-4-27; Published 2009-4-29
Abstract
In many moths, mate-finding communication is mediated by the female sex pheromones. Since differentiation of sex pheromones is often associated with speciation, it is intriguing to know how the changes in female sex pheromone have been tracked by the pheromone recognition system of the males. A male-specific odorant receptor was found to have been conserved through the evolution of sex pheromone communication systems in the genus Ostrinia (Lepidoptera: Crambidae). In an effort to characterize pheromone receptors of O. scapulalis, which uses a mixture of (E)-11- and (Z)-11-tetradecenyl acetates as a sex pheromone, we cloned a gene (OscaOR1) encoding a male-specific odorant receptor. In addition, we cloned a gene of the Or83b family (OscaOR2). Functional assays using Xenopus oocytes co-expressing OscaOR1 and OscaOR2 have shown that OscaOR1 is, unexpectedly, a receptor of (E)-11-tetradecenol (E11-14:OH), a single pheromone component of a congener O. latipennis. Subsequent studies on O. latipennis showed that this species indeed has a gene orthologous to OscaOR1 (OlatOR1), a functional assay of which confirmed it to be a gene encoding the receptor of E11-14:OH. Furthermore, investigations of six other Ostrinia species have revealed that all of them have a gene orthologous to OscaOR1, although none of these species, except O. ovalipennis, a species most closely related to O. latipennis, uses E11-14:OH as the pheromone component. The present findings suggest that the male-specific receptor of E11-14:OH was acquired before the divergence of the genus Ostrinia, and functionally retained through the evolution of this genus.
Keywords: odorant receptor, pheromone receptor, Ostrinia, (E)-11-tetradecenol, functional assay
Introduction
In many moths, mate-finding communication is mediated by the sex pheromones produced by females, which are usually a blend of a few compounds [1]. The species specificity of pheromones is conferred by the combination and blend ratio of components, and the males' response is tuned to the specific blend for ensuring correct finding of conspecific females. Since differentiation of sex pheromones is often associated with speciation, it is intriguing to know how the changes in female sex pheromone have been tracked by the pheromone recognition system of the males.
Moths of the genus Ostrinia (Lepidoptera: Crambidae), which show distinct species-specific differentiation of the sex pheromone communication system [2], are good materials for studying the evolution of the pheromone recognition system. Among the 21 species of the genus Ostrinia recorded worldwide [3, 4], the sex pheromones of nine species (O. nubilalis, O. scapulalis, O. furnacalis, O. orientalis, O. zaguliaevi, O. zealis, O. palustralis, O. latipennis, and O. ovalipennis) have been characterized to date. The following six compounds have been found as the components of sex pheromones produced by females; Z9-14:OAc, E11-14:OAc, Z11-14:OAc, E12-14:OAc, Z12-14:OAc and E11-14:OH (Fig. 1) [2, 5-7]. The adzuki bean borer O. scapulalis, the species mainly focused on in the present study, uses a mixture of E11-14:OAc and Z11-14:OAc as the sex pheromone [8].
Phylogenetic relationships (left) and sex pheromone blends (right) of Ostrinia species examined in this study. The phylogenetic tree was constructed based on the mitochondrial COII gene sequences. The numbers near branches indicate bootstrap values. The size of circles represents rough blend ratio, and × denotes that the compound works as behavioral antagonist. * not tested for behavioral antagonism.
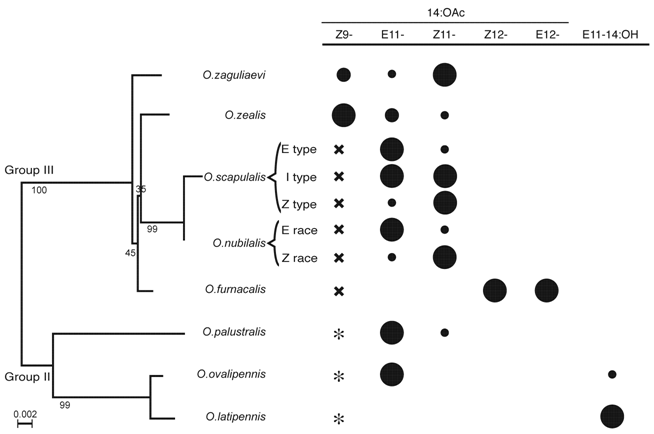
To detect and track the sex pheromones emitted by the conspecific females, males should have a system that can discriminate pheromone components and their ratios. In this system, pheromone receptors in the olfactory sensilla of male moths are likely to play an important role in the specific recognition of pheromone molecules. Recently, information on the molecular characteristics of pheromone receptors in moths, although some may be more appropriately called “male-specific odorant receptors”, has amassed rapidly. Two sex pheromone receptors (BmOR1 and BmOR3) have been characterized in the silkmoth Bombyx mori [9-12], and three pheromone receptors (HR13, HR14 and HR16) have been characterized in the tobacco budworm Heliothis virescens [13-15]. More recently, pheromone receptors were characterized in the diamondback moth Plutella xylostella (PxOR1), the oriental armyworm Mythimna separata (MsOR1), and the cotton caterpillar Diaphania indica (DiOR1) [16].
Along with the characterization of pheromone receptors, the role of Or83b family proteins, co-expressed with odorant receptors (ORs) in most olfactory receptor neurons (ORNs), has been clarified [17-19]. Or83b is considered to act as a chaperone protein in the transport of ORs to the cell membrane of ORNs [20]. Furthermore, Or83b has been recently shown to form a heterodimer with OR, which configures a ligand-gated cation channel [21, 22].
In the present study, as a first step to understanding the evolution of the pheromone reception system in the genus Ostrinia, we tried to clone pheromone receptor genes and an Or83b homologue from the male antennae of O. scapulalis. Since a functional assay of the cloned receptor gene (OscaOR1) revealed that this gene, unexpectedly, encodes a receptor of a pheromone component used in a congener, O. latipennis, but not used in O. scapulalis, we subsequently surveyed homologous genes in O. latipennis and six other congeners, in order to examine the possibility that this receptor has been retained through the evolution of the genus Ostrinia.
Materials and Methods
Insects
Seven species of Ostrinia were reared according to previous reports [6, 23-26]. One-day-old adults and the 5th instar larvae were used in the present study.
RNA extraction and cDNA synthesis
Total RNA was isolated from the antennae, legs, proboscis, testes, head, midgut and fat body of an individual of Ostrinia using an RNeasy Micro Kit (Qiagen). The first-strand cDNA was synthesized with a Takara RNA PCR kit (AMV) Ver.3.0 or a PrimeScript 1st strand cDNA Synthesis kit (Takara Bio Inc.). The quality of the cDNA was verified by testing PCR amplification of the beta-actin gene using the primers sBACT, AACTGGGATGACATGGAGAAGATCTGGC, and aBACT, GAGATCCACATCTGCTGGAAGGTGGACA [34].
RT-PCR and cloning (OscaORs)
Fragments of cDNA encoding odorant receptors of O. scapulalis (OscaORs) were amplified from the first-strand cDNA, which was prepared from the antennae of an E-type male moth of O. scapulalis, using Ex Taq DNA polymerase (Takara-bio) and the following degenerate primers designed based on the consensus sequences of BmORs and HRs: sOR2 (TGYGARCARYTNCARCA), aOR2 (ACCATRAARTANGTNAC), sORA (TGGGSNCAYYTNAARAT), and aORA (TCCCANGGNARNNNRTANAC). The amplification was performed according to the following program: 2 min at 94°C; 1 min at 94°C, 1 min at 40°C and 1 min at 72°C (40 cycles); 15 min at 72°C. The PCR products were cloned into pGEM-T easy vector (Promega) and transformed into Escherichia coli JM109 cells. The sequence of the cloned cDNA was determined with an ABI Prism 310 Genetic Analyzer or ABI Prism 3700 Genetic Analyzer (Applied Biosystems).
Tissue specificity of OscaORs
To investigate the expression of OscaOR1 and OscaOR2 in different tissues, the following primer pairs specific for each OscaOR were used to amplify cDNA from each tissue: OR1 (CAAAAATGGACCCTGAATCG and TCCTTTAGGGTCTCGCTCTG) and OR2 (CCTTCGAGGAGCTGGTGGTAGA and GTGAAGAACTTGGCCCCAGAGA). Negative control experiments were run using samples prepared without reverse transcriptase.
5'- and 3'-RACE
RACE reactions were performed using a CapFishing Full-length cDNA Premix Kit (Seegene Inc.) following the manufacturer's instructions. The 5'-RACE antisense primers and 3'-RACE sense primers were as follows: aOR1-5' (TCCTTTAGGGTCTCGCTCTG), sOR1-3' (CGCAGCCTTGACAGTAACTC), aOR2-5' (CCCATGGATGCCACTAATCTC), and sOR2-3' (ACGTGTACGCGTTCAGCACAAT).
RT-PCR and cloning (ORs of other Ostrinia spp.)
The cDNAs encoding ORs were amplified from the first-strand cDNA prepared from adult male antennae of Ostrinia and primers specific for OR1 and OR2 designed based on the sequences of OscaORs: OR1 (ATGTTTAAAATTGAAAATCA and TTAGCTGAACGTTCGCAAGAACATGAAGTA), OR2 (ATGACCAAAGTGAAAGCT and CTACTTCAGTTGTACCAAAACCATGAAATAAGTCAC). To investigate the expression of ORs in adult antennae, the following primer pairs specific for each OR were used to amplify cDNA (OR1: CAAAAATGGACCCTGAATCG and TCCTTTAGGGTCTCGCTCTG; OR2: CCTTCGAGGAGCTGGTGGTAGA and GTGAAGAACTTGGCCCCAGAGA).
Phylogenetic analysis
The amino acid sequences were aligned using Clustal X [28]. The phylogeny tree was constructed and evaluated by the bootstrap test with 100 re-samplings using the software PHYLIP 3.66 [29].
In situ hybridization
The DNA fragment of OscaORs was subcloned into pGEM-T easy vector (Promega) and used for the generation of sense or antisense RNA probes. Digoxigenin-labeled-RNA probes were prepared with the DIG RNA labeling Mix (Roche). Paraffin-embedded blocks and sections of O. scapulalis for in situ hybridization were obtained by conventional means. Tissue sections were de-waxed with xylene, and re-hydrated through an ethanol series and PBS. The sections were treated for hybridization according to Hoshino et al [30]. Anti-digoxigenin antibody labeled with alkaline phosphatase and BM purple AP substrate (Roche) were used to visualize the signal. The sections were counterstained with Kernechtrot stain solution (Muto), dehydrated, and then mounted with Malinol (Muto).
Chemicals
Z9-14:OAc and Z11-14:OAc were purchased from Sigma Chemicals Co. (St. Louis, MO). E11-14:OAc, E12-14:OAc, Z12-14:OAc and E11-14:OH were obtained from Pherobank (Wageningen, The Netherlands). 14:OAc, Z11-16:OAc and Z11-16:Ald were purchased from Shin-Etsu Chemical Co., Ltd. (Tokyo, Japan). Stock solutions of chemicals (300 mM) were prepared in DMSO and stored at -20°C. Prior to experiments, the stock solution was diluted with bath solution (88 mM NaCl, 1 mM KCl, 0.3 mM Ca(NO3)2, 0.4 mM CaCl2, 0.8 mM MgSO4, 2.4 mM NaHCO3, and 15 mM HEPES, pH 7.6). Bath solution containing 0.1% DMSO was used as a negative control.
Protein expression in Xenopus laevis oocytes and electrophysiological recording
We used the Xenopus oocyte system, which can test the function of putative odorant receptors with high sensitivity [ 9, 16]. Stage V to VII oocytes were treated with 2 mg/ml of collagenase S-1 (Nitta gelatin) in Ca2+-free saline solution (82.5 mM NaCl, 2 mM KCl, 1 mM MgCl2, and 5 mM HEPES, pH 7.5) for 1 to 2 h at room temperature. Oocytes were then microinjected with 25 ng of cRNA for a conventional OR (OscaOR1 and OlatOR1) and 25 ng of cRNA for the Or83b family (OscaOR2 and OlatOR2). cRNA was synthesized from linearized modified pSPUTK [38]. Injected oocytes were incubated for 3 days at 18°C in bath solution supplemented with 10 µg/ml of penicillin and streptomycin. Whole-cell currents were recorded with a two-electrode voltage-clamp filled with 3 M KCl, amplified with an OC-725C amplifier (Warner Instruments), low-pass filtered at 50 Hz and digitized at 1 kHz. The inward current was monitored at a holding potential of -80 mV. Pheromones were delivered through the superfusing bath solution via a silicon tube connected to computer-driven electric valves. Data acquisition and analysis were carried out using Digidata1322A and pCLAMP software.
Results
cDNA cloning of a putative sex pheromone receptor of O. scapulalis
A full-length cDNA of a putative sex pheromone receptor gene (OscaOR1) was cloned (Fig. 2A). The deduced amino acid sequence of OscaOR1 showed 34-41% overall identity to male-specific BmORs of B. mori and HRs of H. virescens, the C-terminal being the most conserved region (Fig. 2B). Subsequently, a cDNA (OscaOR2) that encodes an Or83b family protein was cloned (Fig. 3A). The deduced amino acid sequence of OscaOR2 showed a high degree of identity to Or83b family proteins of various insect species belonging to different orders (Fig. 3B).
Nucleotide and deduced amino acid sequences of OscaOR1. A, full nucleotide sequences and deduced amino acid sequence of OscaOR1. Arrows indicate primers using RT-PCR. B, aligned amino acid sequences of OscaOR1 and sex pheromone receptors of Bombyx mori and Heliothis virescens.

Nucleotide and deduced amino acid sequences of OscaOR2. A, full nucleotide sequence and deduced amino acid sequence of OscaOR2. Arrows indicate primers using RT-PCR. B, Aligned amino acid sequences of OscaOR2 and Or83b family proteins. OscaOR2 (Ostrinia scapulalis, present study), DmOr83b (Drosophila melanogaster), AgOR7 (Anopheles gambiae), HvOR2 (H. virescens), BmOR2 (B. mori), TcOr1 (Tribolium castaneum), and AmOr2 (Apis mellifera).

Expression of OscaOR1 and OscaOR2 in different tissues
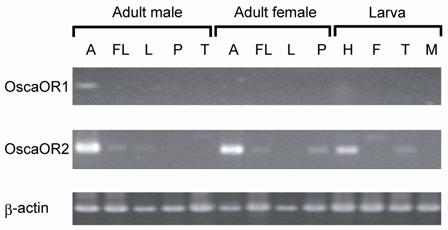
The expression of OscaOR1 and OscaOR2 in tissues other than adult male antennae was examined by RT-PCR (Fig.4). The mRNA of OscaOR1 was not expressed in any tissues examined (adult female antennae, the legs, testes, proboscis of an adult moth, and the head, midgut, fat body, and testes of a larva), demonstrating that the expression of this gene is specific to the male antenna. On the other hand, the mRNA of OscaOR2 was expressed in the larval head, adult foreleg and adult female proboscis, in addition to the antennae of adult females (Fig.4). This finding is in accord with the recent consensus that Or83b family proteins are expressed widely in chemosensory organs [19, 32-34].
Expression of OR1 and OR2 genes in different tissues. Expression of OscaOR1 and OscaOR2 in different tissues was tested by RT-PCR using specific primers for each gene. A, antenna; FL, foreleg; L, midleg and hindleg; P, proboscis; T, testis; H, head; F, fat body; M, midgut.
In situ hybridization
In the longitudinal sections of antennae, many cell bodies at the base of sensilla trichodea or basiconica were stained with the antisense probes for OscaOR1 and OscaOR2 (Fig.5A, C, D, F), although the number of cells stained differed with the probe used. In contrast, no cell bodies at the base of sensilla coeloconica were stained with either antisense probe (Fig.5C).
Expression of OscaOR1 and OscaOR2 genes in adult male antenna. Longitudinal sections of antennae were hybridized with antisense or sense probes for OscaOR2 (A, B and C). Longitudinal sections of antennae were hybridized with antisense or sense probes for OscaOR1 (D, E and F). Magnification of the boxed area in A (C) and D (F). Arrowheads indicate the cell bodies stained. Arrows indicate sensilla coeloconica. Scale bar in A = 10 µm (the same scale applies to A, B, D and E). Scale bar in C = 2 µm (the same scale applies to C and F).
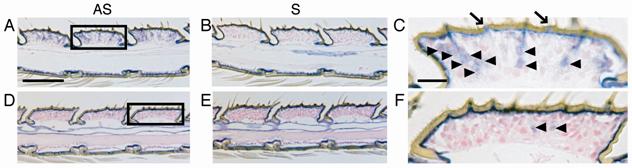
Given that the Or83b family proteins are broadly expressed in ORNs [18, 32, 33, 35, 36], the cells stained with the antisense probe for OscaOR2 (Or83b homologue) were considered to represent most ORNs. Accordingly, the percentage of ORNs expressing OscaOR1 was estimated to be about 20%, since the numbers of cells stained with the antisense probes for OscaOR1 and OscaOR2 were 1-3 (mean = 1.8; Fig.5D, F) and 9-14 (mean = 10.1; Fig.5C) per antennal segment in the longitudinal sections, respectively. Likewise, given that the number of sensilla trichodea or basiconica was 4-5 per antennal segment (Fig.5), 2-3 ORNs were estimated to be associated with each sensillum. This estimated number of ORNs per sensillum was well consistent with the results of an electrophysiological study using O. nubilalis [37].
Functional assay of OscaOR1
To check if the OscaOR1 protein is a sex pheromone receptor, we examined electrophysiological responses of Xenopus oocytes co-expressing OscaOR1 and OscaOR2 (OscaOR1/OscaOR2 cell) to the 6 sex pheromone components used in the genus Ostrinia and three pheromone analogs. We confirmed beforehand that adult male antennae of O. scapulalis respond to these 9 compounds using electroantennography (data not shown).
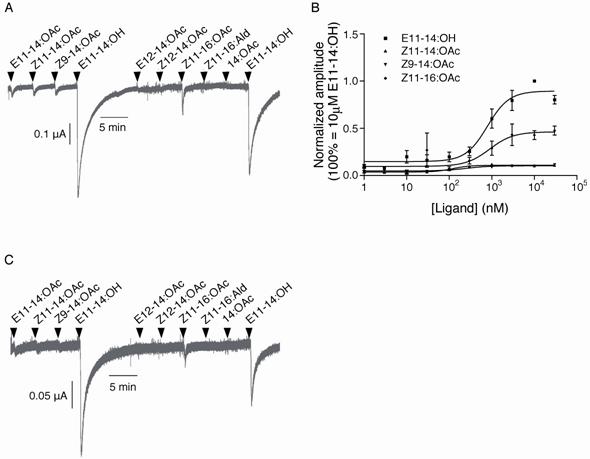
OscaOR1/OscaOR2 cells showed a large response to E11-14:OH and a small response to Z11-16:OAc (Fig.6A), while the control cells without artificially expressed proteins showed no responses (data not shown). The responses of OscaOR1/OscaOR2 cells to E11-14:OH and Z11-16:OAc were dose-dependent (Fig.6B). E11-14:OH is not a sex pheromone component of O. scapulalis, but a single pheromone component of O. latipennis and a minor pheromone component of O. ovalipennis [6, 7]. Unexpectedly, OscaOR1/OscaOR2 cells did not show significant responses to E11-14:OAc and Z11-14:OAc, the sex pheromone components of O. scapulalis.
Responses of oocytes co-expressing OR1 and OR2 to the pheromone components and analogs. A, responses of oocytes co-expressing OscaOR1/OscaOR2. B, dose-response curves of four compounds to which the oocytes showed significant responses. C, responses of oocytes co-expressing OlatOR1 and OlatOR2, orthologs of OscaOR1 and OscaOR2 in O. latipennis, respectively. The concentrations of pheromone components in A and C were10 µM.
cDNA cloning of OR1s of the genus Ostrinia
We were able to clone homologues of OscaOR1 from all the Ostrinia species examined (Fig.7). A single OscaOR1 homologue was obtained from each species: OlatOR1 (O. latipennis), OfurOR1 (O. furnacalis), OzagOR1 (O. zaguliaevi), OzeaOR1 (O. zealis), OpalOR1 (O. palustralis), OovaOR1 (O. ovalipennis), and OnubOR1 (O. nubilalis). High degrees of identity (93-99%) were observed between the deduced amino acid sequences of these OR1s (Fig.7). In the phylogenetic tree of odorant receptor proteins, OR1s of the genus Ostrinia formed a clade, suggesting that these genes are orthologous (OR1 group, Fig.8). Moreover, the OR1 group was included in a single lineage of the sex pheromone receptor subfamily.
Aligned amino acid sequences of OR1 orthologs in the genus Ostrinia. The gray boxes show positions where a substitution of the amino acid residue was found between OscaOR1 and OlatOR1.

Phylogenetic tree of odorant receptor proteins of Ostrinia and other lepidopteran species. The numbers near branches indicate bootstrap values. Values of < 70% are not shown.

Functional assay of OlatOR1
In order to verify the function of OlatOR1 as an odorant receptor of E11-14:OH, we additionally cloned an Or83b homologue from O. latipennis (OlatOR2), and examined responses of oocytes co-expressing OlatOR1 and OlatOR2 (OlatOR1/OlatOR2 cell) to the 9 compounds (Fig.6C). OlatOR1/OlatOR2 cells were found to show exactly the same response pattern as OscaOR1/OscaOR2 cells.
Discussion
Based on the morphology of the male genitalia, Ostrinia species found in Japan and the European corn borer O. nubilalis are classified into two groups, i.e., a simple or bifid uncus group (Group II; O. palustralis, O. latipennis, and O. ovalipennis) and a trilobed uncus group (Group III; O. nubilalis, O. scapulalis, O. furnacalis, O. orientalis, O. zaguliaevi, and O. zealis) [3]. This grouping is also supported by molecular phylogenetic analyses of the mitochondrial cytochrome oxidase subunit II (COII) gene (Fig. 1) [38]. In the phylogenetic tree of odorant receptor proteins, the OR1 group was divided into two subgroups, the members of which perfectly matched with those of groups II and III, respectively (Fig.8). These findings suggest that speciation within the genus Ostrinia occurred after the acquirement of OR1.
The OR1s of O. scapulalis and O. latipennis, OscaOR1 and OlatOR1, strongly responded to E11-14:OH, indicating that OscaOR1 and OlatOR1 are indeed the receptor proteins of E11-14:OH. Since E11-14:OH is a single sex pheromone component of O. latipennis [6], it is concluded that OlatOR1 functions as the sex pheromone receptor of O. latipennis. Although E11-14:OH is not a pheromone component of O. scapulalis, OscaOR1 also acts as the receptor of E11-14:OH because electroantennographic recordings showed that the male antennae of O. scapulalis respond to E11-14:OH (data not shown). This ability of O. scapulalis to detect E11-14:OH, a pheromone component of a congener, may suggest that change(s) in the neural circuit in the peripheral or central nervous system, rather than change(s) in the pheromone receptor, is responsible for the changes in the pheromone recognition system of Ostrinia. In this regard, E11-14:OH might have an unknown function in the pheromone communication system of O. scapulalis, as discussed below.
Within the genus, O. scapulalis and O. latipennis are most distantly related [2, 3]. Very high degrees of identity between the amino acid sequences of OR1s in Ostrinia species including O. scapulalis and O. latipennis suggest that all OR1s in this genus are receptors of E11-14:OH. It is curious that the receptor of E11-14:OH is conserved in all Ostrinia species examined because most of them do not use E11-14:OH as the pheromone component. Regarding the significance of OR1s in this genus, two possibilities can be considered. One is that E11-14:OH has an unknown role in the sex pheromone communication systems of the genus Ostrinia. For instance, this compound might serve as an antagonist in species other than O. latipennis and O. ovalipennis, although no one has systematically tested this possibility to date. The other possibility is that the common ancestor of the genus Ostrinia used E11-14:OH as a pheromone component, and despite that many species stopped using this component in the course of speciation, sufficient time for mutational loss of the receptor function may not have passed yet.
In OscaOR1 and OlatOR1, substitutions of amino acid residues were found at 26 positions (Fig.7). Because OscaOR1 and OlatOR1 showed the same pattern of responses to the nine compounds tested, these 26 amino acid positions are not essential for the function of this receptor, suggesting that amino acids at these positions are not involved in the formation of the binding site of E11-14:OH or formation of the ion channel. It is difficult to tell whether the function of OR1s has been positively retained through evolution or not, since the low amino acid substitution rates found in OR1s are natural given that species in the genus Ostrinia diverged only within 0.4 million years [38].
OscaOR1 was expressed in about 20% of ORNs in the antennae of male O. scapulalis. Male-specific odorant receptors other than OscaOR1, which remain to be characterized, are expected to appear in the remaining 80% of ORNs. The receptors of E11-14:OAc and Z11-14:OAc, the sex pheromone components of O. scapulalis, are likely to be included in yet unidentified receptors. All Ostrinia species except for O. latipennis and O. furnacalis use E11-14:OAc and/or Z11-14:OAc as sex pheromone components [2]. Hence, the use of E11-14:OAc and Z11-14:OAc as the sex pheromone components in Ostrinia, and accordingly, the acquisition of receptors of E11-14:OAc and Z11-14:OAc are likely to ascend to the divergence of the genus. We therefore suspect that the receptors of E11-14:OAc and Z11-14:OAc are also conserved in the genus Ostrinia. It is intriguing to know the repertory of pheromone receptors in the ancestors of Ostrinia, because it might have delimited the possible variation in the pheromone communication system in this genus.
Acknowledgements
We thank Drs. Ryo Nakano, Takeshi Fujii and Sugihiko Hoshizaki for help in insect rearing and useful discussions. This work was supported by a Grant-in-aid for scientific research No. 19208005 (to Y. I.) from the Japan Society for the Promotion of Science.
The abbreviations used are: 14:OAc, tetradecyl acetate; Z9-14:OAc, (Z)-9-tetradecenyl acetate; E11-14:OAc, (E)-11-tetradecenyl acetate; Z11-14:OAc, (Z)-11-tetradecenyl acetate; E12-14:OAc, (E)-12-tetradecenyl acetate; Z12-14:OAc, (Z)-12-tetradecenyl acetate; E11-14:OH, (E)-11-tetradecenol; Z11-16:OAc, (Z)-11-hexadecenyl acetate; Z11-16:Ald, (Z)-11-hexadecenal; HEPES, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; DMSO, dimethyl sulfoxide; RACE, rapid amplification of cDNA ends; RT-PCR, reverse transcription- polymerase chain reaction. The nucleotide sequences reported in this paper has been submitted to the DDBJ/GenBankTM/EBI Data Bank with accession number(s) AB467318 (OscaOR2), AB467319 (OlatOR2), AB467320 (OscaOR1), AB467321 (OzagOR1), AB467322 (OzeaOR1), AB467323 (OpalOR1), AB467324 (OovaOR1), AB467325 (OnubOR1), AB467326 (OlatOR1), and AB467327 (OfurOR1).
Conflict of Interests
The authors have declared that no conflict of interest exists.
References
1. The Pherolist. Witzgall P, Lindblom T, Bengtsson M, Toth M. http://www-pherolist.slu.se
2. Ishikawa Y, Takanashi T, Kim C, Hoshizaki S, Tatsuki S, Huang YP. Ostrinia spp. in Japan: their host plants and sex pheromones. Entomol Exp Appl. 1999;91:237-44
3. Mutuura A, Munroe E. Taxonomy and distribution of corn borer and allied species - genus Ostrinia (Lepidoptera-Pyralidae). Mem. Entomol. Soc. Can. 1970;71:1-112
4. Ohno S. A new knotweed-boring species of the genus Ostrinia Hubner (Lepidoptera: Crambidae) from Japan. Entomol. Sci. 2003;6:77-83
5. Roelofs WL, Du JW, Tang XH, Robbins PS, Eckenrode CJ. 3 European corn-borer populations in New York based on sex-pheromones and voltinis. J. Chem. Ecol. 1985;11:829-36
6. Takanashi T, Ohno S, Huang YP, Tatsuki S, Honda H, Ishikawa Y. A sex pheromone component novel to Ostrinia identified from Ostrinia latipennis (Lepidoptera: Crambidae). Chemoecology. 2000;10:143-7
7. Ohno S. Systematics of Ostrinia latipennis species group based on morphology, DNA and sex pheromones. PhD dissertation. The university of Tokyo, Japan (in Japanese). 2000
8. Takanashi T, Huang YP, Takahasi KR, Hoshizaki S, Tatsuki S, Ishikawa Y. Genetic analysis and population survey of sex pheromone variation in the adzuki bean borer moth, Ostrinia scapulalis. Biol. J. Linn. Soc. 2005;84:143-60
9. Sakurai T, Nakagawa T, Mitsuno H, Mori H, Endo Y, Tanoue S, Yasukochi Y, Touhara K, Nishioka T. Identification and functional characterization of a sex pheromone receptor in the silkmoth Bombyx mori. Proc. Natl. Acad. Sci. U. S. A. 2004;101:16653-8
10. Krieger J, Grosse-Wilde E, Gohl T, Breer H. Candidate pheromone receptors of the silkmoth Bombyx mori. Eur. J. Neurosci. 2005;21:2167-76
11. Nakagawa T, Sakurai T, Nishioka T, Touhara K. Insect sex-pheromone signals mediated by specific combinations of olfactory receptors. Science. 2005;307:1638-42
12. Syed Z, Ishida Y, Taylor K, Kimbrell DA, Leal WS. Pheromone reception in fruit flies expressing a moth's odorant receptor. Proc. Natl. Acad. Sci. U. S. A. 2006;103:16538-43
13. Krieger J, Grosse-Wilde E, Gohl T, Dewer YME, Raming K, Breer H. Genes encoding candidate pheromone receptors in a moth (Heliothis virescens). Proc. Natl. Acad. Sci. U. S. A. 2004;101:11845-50
14. Gohl T, Krieger J. Immunolocalization of a candidate pheromone receptor in the antenna of the male moth, Heliothis virescens. Invertebr. Neurosci. 2006;6:13-21
15. Kurtovic A, Widmer A, Dickson BJ. A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature. 2007;446:542-6
16. Mitsuno H, Sakurai T, Murai M, Yasuda T, Kugimiya S, Ozawa R, Toyohara H, Takabayashi J, Miyoshi H, Nishioka T. Identification of receptors of main sex-pheromone components of three Lepidopteran species. Eur. J. Neurosci. 2008;28:893-902
17. Vosshall LB, Wong AM, Axel R. An olfactory sensory map in the fly brain. Cell. 2000;102:147-59
18. Krieger J, Klink O, Mohl C, Raming K, Breer H. A candidate olfactory receptor subtype highly conserved across different insect orders. J. Comp. Physiol. A. 2003;189:519-26
19. Larsson MC, Domingos AI, Jones WD, Chiappe ME, Amrein H, Vosshall LB. Or83b encodes a broadly expressed odorant receptor essential for Drosophila olfaction. Neuron. 2004;43:703-14
20. Benton R, Sachse S, Michnick SW, Vosshall LB. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. Plos Biol. 2006;4:240-57
21. Sato K, Pellegrino M, Nakagawa T, Vosshall LB, Touhara K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature. 2008;452:1002-1006
22. Wicher D, Schafer R, Bauernfeind R, Stensmyr MC, Heller R, Heinemann SH, Hansson BS. Drosophila odorant receptors are both ligand-gated and cyclic-nucleotide-activated cation channels. Nature. 2008;452:1007-1011
23. Huang YP, Tatsuki S, Kim CG, Hoshizaki S, Yoshiyasu Y, Honda H, Ishikawa Y. Identification of sex pheromone of adzuki bean borer, Ostrinia scapulalis. J. Chem. Ecol. 1997;23:2791-802
24. Huang YP, Honda H, Yoshiyasu Y, Hoshizaki S, Tatsuki S, Ishikawa Y. Sex pheromone of the butterbur borer, Ostrinia zaguliaevi. Entomol. Exp. Appl. 1998;89:281-7
25. Ishikawa Y, Takanashi T, Huang Y. Comparative studies on the sex pheromones of Ostrinia spp. in Japan: the burdock borer, Ostrinia zealis. Chemoecology. 1999;9:25-32
26. Fukuzawa M, Tatsuki S, Ishikawa Y. Rearing of Ostrinia palustralis (Lepidoptera: Crambidae) larvae with a switchover of two kinds of artificial diets. Appl. Entomol. Zool. 2004;39:363-6
27. Miura N, Atsumi S, Tabunoki H, Sato R. Expression and localization of three G protein alpha subunits, G(o), G(q), and G(s), in adult antennae of the silkmoth (Bombyx mori). J. Comp. Neurol. 2005;485:143-52
28. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. Clustal W and clustal X version 2.0. Bioinformatics. 2007;23:2947-8
29. Felsenstein J. PHYLIP (Phylogeny Inference Package) version 3.66, 2004.
30. Hoshino M, Sone M, Fukata M, Kurodad S, Kaibuchi K, Nabeshima Y, Hama C. Identification of the stef gene that encodes a novel guanine nucleotide exchange factor specific for Rac1. J. Biol. Chem. 1999;274:17837-44
31. Katada S, Nakagawa T, Kataoka H, Touhara K. Odorant response assays for a heterologously expressed olfactory receptor. Biochem. Biophys. Res. Commun. 2003;305:964-9
32. Melo ACA, Rutzler M, Pitts RJ, Zwiebel LJ. Identification of a chemosensory receptor from the yellow fever mosquito, Aedes aegypti, that is highly conserved and expressed in olfactory and gustatory organs. Chem. Senses. 2004;29:403-10
33. Pitts RJ, Fox AN, Zwiebel LJ. A highly conserved candidate chemoreceptor expressed in both olfactory and gustatory tissues in the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. U. S. A. 2004;101:5058-63
34. Xia YF, Zwiebel LJ. Identification and characterization of an odorant receptor from the West Nile Virus mosquito, Culex quinquefasciatus. Insect Biochem. Mol. Biol. 2006;36:169-76
35. Vosshall LB, Amrein H, Morozov PS, Rzhetsky A, Axel R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell. 1999;96:725-36
36. Krieger J, Raming K, Dewer YME, Bette S, Conzelmann S, Breer H. A divergent gene family encoding candidate olfactory receptors of the moth Heliothis virescens. Eur. J. Neurosci. 2002;16:619-28
37. Hallberg E, Hansson BS, Steinbrecht RA. Morphological- characteristics of antennal sensilla in the European cornborer Ostrinia-nubilalis (Lepidoptera, Pyralidae). Tissue & Cell. 1994;26:489-502
38. Kim CG, Hoshizaki S, Huang YP, Tatsuki S, Ishikawa Y. Usefulness of mitochondrial COII gene sequences in examining phylogenetic relationships in the Asian corn borer, Ostrinia furnacalis, and allied species (Lepidoptera: Pyralidae). Appl. Entomol. Zool. 1999;34:405-12
Author contact
![]() Correspondence to: Dr. Yukio Ishikawa, Department of Agricultural and Environmental Biology, Graduate School of Agricultural and Life Sciences, University of Tokyo, Tokyo 113-8657, Japan; Tel and Fax: 81-3-5841-5061; E-mail: ayuckyecc.u-tokyo.ac.jp (Y. Ishikawa)
Correspondence to: Dr. Yukio Ishikawa, Department of Agricultural and Environmental Biology, Graduate School of Agricultural and Life Sciences, University of Tokyo, Tokyo 113-8657, Japan; Tel and Fax: 81-3-5841-5061; E-mail: ayuckyecc.u-tokyo.ac.jp (Y. Ishikawa)