Impact Factor ISSN: 1449-2288
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Issue 9; 2026
- Issue 8; 2026
- Volume 22; 2026
- Past Issues
- Advance Articles
- Editorial Board
- Cover Images
- Index & Coverage
- Cover Suggestion
- Special Issues
Introduction
M7G regulators and their roles...
The roles of m7G modification in...
M7G regulate TIME by affecting...
M7G modification influence the...
Conclusion and prospects
Abbreviations
Acknowledgements
References

 Global reach, higher impact
Global reach, higher impactInt J Biol Sci 2024; 20(4):1238-1255. doi:10.7150/ijbs.90382 This issue Cite
Review
M7G-related tumor immunity: novel insights of RNA modification and potential therapeutic targets
Mengzhen Han1,2, Qibo Huang1,2, Xinxin Li1,2, Xiaoping Chen1,2, He Zhu1,2 ![]() , Yonglong Pan1,2
, Yonglong Pan1,2 ![]() , Bixiang Zhang1,2
, Bixiang Zhang1,2 ![]()
1. Hepatic Surgery Center, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei 430030, China.
2. Hubei Key Laboratory of Hepato-Pancreato-Biliary Diseases, Wuhan, Hubei 430030, China.
Received 2023-9-21; Accepted 2024-1-9; Published 2024-1-27
Abstract

RNA modifications play a pivotal role in regulating cellular biology by exerting influence over distribution features and molecular functions at the post-transcriptional level. Among these modifications, N7-methylguanosine (m7G) stands out as one of the most prevalent. Over recent years, significant attention has been directed towards understanding the implications of m7G modification. This modification is present in diverse RNA molecules, including transfer RNAs, messenger RNAs, ribosomal RNAs, and other noncoding RNAs. Its regulation occurs through a series of specific methyltransferases and m7G-binding proteins. Notably, m7G modification has been implicated in various diseases, prominently across multiple cancer types. Earlier studies have elucidated the significance of m7G modification in the context of immune biology regulation within the tumor microenvironment. This comprehensive review culminates in a synthesis of findings related to the modulation of immune cells infiltration, encompassing T cells, B cells, and various innate immune cells, all orchestrated by m7G modification. Furthermore, the interplay between m7G modification and its regulatory proteins can profoundly affect the efficacy of diverse adjuvant therapeutics, thereby potentially serving as a pivotal biomarker and therapeutic target for combinatory interventions in diverse cancer types.
Keywords: N7-methylguanosine (m7G), RNA modification, Tumor immunity, Adjuvant therapy, Biomarker
Introduction
The progression of detection technologies in recent decades has led to a heightened emphasis on investigating the biological roles of RNA modifications [1]. RNA modifications are chemical alterations that take place within the ribose and nucleobase constituents. These alterations significantly impact the structural conformation and functional properties of RNA molecules [2-4]. There are more than 150 types of identified RNA modifications, including pseudouridine (ψ), N6-methyladenosine (m6A), N1-methyladenosine (m1A), 5-methylcytosine (m5C), N7-methylguanosine (m7G), N4 -acetylcytosine (ac4C), uridylation, adenosine-to-inosine (A-to-I) RNA editing, etc [5,6]. Among these different RNA modifications, m6A is most commonly found in messenger RNA (mRNA) modification, m5C in transfer RNA (tRNA) can influence its stability and translational fidelity, while ψ mainly present in non-coding RNA (ncRNA) like ribosomal RNA (rRNA) and small nuclear RNAs (snRNAs) [7-9]. More recently, researchers also reported several RNA modifications in microRNAs (miRNAs), long noncoding RNAs (lncRNAs) and small nucleolar RNAs (snoRNAs) [10,11]. Previous studies have focused on the m6A modification and its role in diverse biological processes [12]. However, further research is necessary to explore additional non-m6A chemical modifications in various types of RNAs.
M7G modification is a methylation of the seventh N on RNA guanylate which can introduce positively charged or amphiphilic ions into nucleobases [13,14]. The m7G modification can be categorized into two distinct types based on the position of the methylated guanylate: m7G-caps modification and internal m7G modification. Initially, the modification of m7G-caps was detected on the 5′ caps of eukaryotic mRNAs. These m7G-caps play a pivotal role in shielding mRNA from degradation while facilitating essential caps-associated mRNA processes, such as splicing, nuclear export, and translation [15,16]. Furthermore, m7G methylation of internal positions was demonstrated in mRNAs, tRNAs and rRNAs, which could also influence the distribution, structure and function of them [17,18]. With the current advancements in methods for detecting m7G-methylated RNAs and high-throughput technologies, an increasing number of methylation sites and diverse types of non-coding RNAs (ncRNAs) have been found to be associated with m7G modification [19,20]. Moreover, earlier studies have proposed that disruption in m7G methylation may link to a spectrum of ailments, encompassing cardiovascular diseases, neurological disorders, and various types of cancers [13,15,21].
Cancer stands as a significant public health challenge and represents the second leading cause of global mortality. The reprograming of immune system and evasion of cancer cells from immune destruction are important hallmarks of cancer, which have been sufficiently validated in past decades [22,23]. Recent researches demonstrated that m7G methylation could regulate immune cells activation and infiltration, which were related to immune surveillance and immune evasion [13,24]. Furthermore, there is a growing focus on tumor-targeted therapies, particularly within the realm of cancer immunotherapy. Notably, the influence of m7G methylation extends to the efficacy of such therapeutic interventions [5]. This comprehensive review primarily aims to elucidate the role of RNA m7G modification in the context of tumor immunity and targeted therapeutic approaches. The insights garnered herein hold the potential to furnish promising strategies for advancing cancer therapy.
M7G regulators and their roles in cancers
Eukaryotic RNA methylation is typically governed by three distinct categories of proteins: methyltransferases (referred to as "writers"), methylation binding proteins (commonly known as "readers"), and demethylases (known as "erasers") [25,26]. Previous research has substantiated the presence of various regulators associated with m7G modification. In yeast, it has been reported that Abd1 functions as a writer responsible for m7G capping of mRNA at the 5' end, while the Trm8/Trm82 complex is responsible for internal m7G modification of tRNAs [18,27]. In humans, regulators of m7G modification encompass a writer complex that includes RNMT/RAM, METTL1/WDR4, and WBSCR22/TRMT112, along with the reader complexes cap-binding complex (CBC) and eukaryotic translation initiation factor 4E (EIF4E) that recognize m7G caps [28,29]. Furthermore, recent investigations have made significant strides in identifying internal m7G binding proteins, which may serve as novel readers of m7G modifications. This paper focuses on elucidating the functions of these regulators in the context of cancer. Figure 1 and 2 depict the relevant regulators and mechanisms associated with m7G capping and internal m7G modification.
Biological mechanism and associated regulators of m7G-cap modification. M7G-cap modification occur in 5′ terminal guanosine of mRNAs and some ncRNAs such as miRNAs, lncRNAs and snoRNAs. M7G capping is an important post-transcriptional process which can influence transcripts splicing, maturation, nuclear export and translation. RNMT/RAM complex is the confirmed writer in m7G-cap modification. CBC, EIF4E and LARP1 are readers which can bind to the m7G caps.
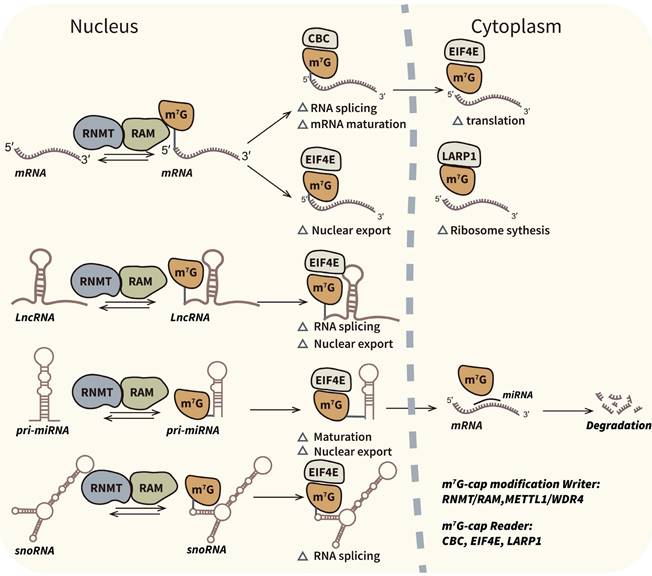
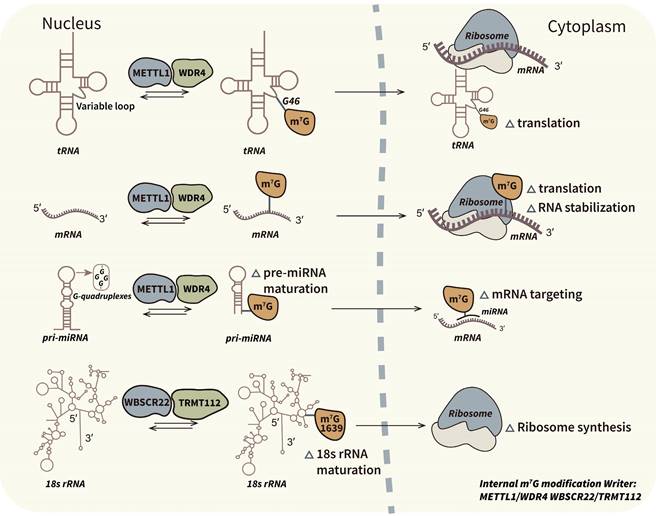
Biological mechanism and associated regulators of internal m7G modification. M7G modification has been demonstrated at internal positions of different types of transcripts. Internal m7G modification plays pivotal roles in RNA stabilization, translation initiation and ribosome synthesis. METTL1/WDR4 complex is the most studied internal m7G writer which catalyze methylation of tRNAs, mRNAs and pri-miRNAs. And WBSCR22/TRMT112 complex is the specific m7G writer of rRNAs.
Writers
RNMT/RAM
M7G-cap modification is a tightly regulated process occurred in 5′ terminal guanosine of mRNAs and certain ncRNAs, which is required for transcripts splicing, cleavage and polyadenylation (CPA), nuclear export and translation initiation in eukaryotes [30]. In the beginning, RNA guanine-7 methyltransferase (RNMT) was considered to be the monomer methyltransferase to catalyze 5' cap methylation by utilizing methyl of S-adenosyl methionine (SAM) [31]. Nevertheless, more recent researches demonstrated that RNMT-Activating Miniprotein (RAM) could bind to RNMT and form an enzyme complex, thereby stabilize and allosterically activate RNMT, as well as increase its affinity for RNA [31-33]. The RNMT/RAM complex is mainly located in nucleus, and the RNAs can directly bind to m7G-cap readers shortly after capping process [34].
Function of RNMT/RAM complex in m7G-cap modification could be regulated by distinct molecules and signaling pathways, which is also associated with various cancers. For example, recent publications suggested that the classical tumorigenic molecule c-MYC could upregulate the m7G-capping process catalyzed by RNMT. In breast cancers, c-MYC was reported to combine with RNMT and enhance the translation of cyclin D1 through the induction of m7G-capping, thus facilitate the malignant transforming of mammary epithelial cells [35]. Similarly, a model of Burkitt's lymphoma showed that c-MYC could recruit RNMT to promoters of WNT pathway genes and promote m7G-capping of these transcripts [36]. Additionally, CDK1 and cyclin B1 could phosphorylate RNMT at Thr77, consequently elevate m7G-cap modification levels and induce cell proliferation [37]. The m7G-cap reader EIF4E were also found in regulation of RNMT, which could serve as co-factor and facilitate the capture of capped RNAs [38].
METTL1/WDR4
The methyltransferase-like 1 (METTL1) and WD repeat domain 4 (WDR4) complex was firstly reported to act as the writer of m7G modification at tRNA variable loop G46[39]. Within this complex, METTL1 recognizes tRNA via its αC and α6 helices, inducing tRNA bending through its N-terminal regions (residues 1-33) while performing catalytic functions. Simultaneously, WDR4 interacts with METTL1, serving as a scaffold to facilitate tRNA binding [40,41]. Earlier investigations have proposed that METTL1 can undergo phosphorylation at Ser27 by AKT and RSK, leading to reduced methyltransferase activity[42]. The m7G modification by the METTL1/WDR4 complex stabilizes tRNA structures, prevents frame shifting, and enhances the translation of mRNAs enriched with m7G-decoded codons [43]. Dysregulation of tRNA m7G modification by METTL1/WDR4 complex and consequent codon-biased translation may contribute to various types of cancers [44,45]. For instance, studies in lung cancer have illustrated that overexpression of METTL1 and WDR4 stabilizes m7G-modified tRNA ValAAC and ProAGG, consequently promoting the translation of genes involved in cell-cycle regulation, thereby influencing tumor progression and prognosis [46]. Similar upregulations of oncogenic translation were also indicated in hepatocellular carcinoma (HCC) and intrahepatic cholangiocarcinoma (ICC) [47,48]. In esophageal squamous cell carcinoma (ESCC), researchers demonstrated that METTL1 and WDR4 could induce the translation of RPTOR and decrease autophagic flux in ESCC cells [49]. Apart from these types of solid tumors, METTL1/WDR4 complex was reported to be involved in oncogenic transformations of acute myelocytic leukemia (AML) in an tRNA m7G dependent manner [50]. These results showed important function of tRNA m7G modification regulated by METTL1/WDR4 complex in cancers.
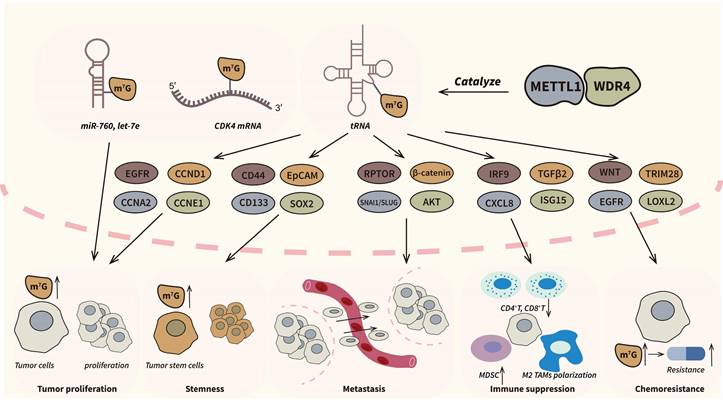
Except for tRNA, METTL1/WDR4 can also catalyze m7G modification of mRNA and miRNA [51]. Transcriptome-wide profiling has unveiled the ability of the METTL1/WDR4 complex to introduce m7G modifications into mRNA molecules, conferring protection against degradation and enhancing their translational efficiency [19]. As internal m7G modification of mRNAs mainly occur in GA-rich context, researchers further indicated that METTL1/WDR4 complex could bind to GA-rich motifs and exert its methyltransferase activity [17,52]. Notably, in prostate cancer, METTL1 stabilizes CDK14 mRNA by catalyzing internal m7G modification, resulting in the upregulation of CDK14, which in turn promotes tumor cell proliferation and invasion [53]. Additionally, Pandolfini et al. firstly reported the m7G modification of miRNA let-7. Their results indicated that METTL1 could facilitates disruption of the secondary structure G-quadruplexes and promote pre-miRNA maturation [54]. Similarly, researches in bladder cancer showed that METTL1 could accelerate tumor progression by overexpressing miR-760 in an m7G-dependent manner [55]. Furthermore, recent studies also reflected the potential m7G modification in lncRNA, which further illustrate the biological function of METTL1/WDR4 complex [56]. A summary of the functions of the METTL1/WDR4 complex in cancers is provided in Figure 3.
The roles of METTL1/WDR4 complex in cancers. METTL1/WDR4 complex is the most extensively researched m7G writer and an important oncogenic factor in many types of cancers. The complex can regulate a series of pathological processes in tumor progression including proliferation, metastasis, stemness, immune suppression and chemoresistance by m7G dependent manners. The dysregulated pathways and genes are showed above.
WBSCR22/TRMT112
The Williams-Beuren syndrome chromosomal region 22 (WBSCR22), also known as metastasis-related methyltransferase 1 (MERM1), was initially identified as an essential pro-metastatic regulator involved in breast cancer metastasis [57]. WBSCR22 functions as a functional homolog of the yeast Bud23 methyltransferase, possessing an SAM binding domain that facilitates the catalysis of 18S rRNA m7G modification at position G1639, thus expediting its maturation [58,59]. These processes play a crucial role in the biogenesis of 40S ribosomal subunits [60]. WBSCR22 has the capacity to interact with its co-factor and activator, the tRNA methyltransferase activator subunit 11-2 (TRMT112), forming a heterodimeric methyltransferase complex responsible for rRNA m7G modification [61]. Previous studies have demonstrated that TRMT112 can stabilize WBSCR22 by inhibiting its Ubiquitin-Proteasome degradation [64]. WBSCR22 and TRMT112 have been implicated in tumor progression and chemoresistance across various cancers. For instance, recent publications in colorectal cancers (CRC) have suggested that the overexpression of WBSCR22 correlates with decreased overall survival and oxaliplatin resistance [62]. Conversely, researchers have also elucidated the tumor-suppressive functions of WBSCR22 and TRMT112 in pancreatic cancers [63]. Nevertheless, the specific mechanisms underlying rRNA m7G modification by the WBSCR22/TRMT112 complex remain underexplored in the context of cancer, and further investigation is warranted to explore other potential m7G writers.
Readers and erasers
In the realm of RNA methylation, erasers possess the ability to reversibly remove methyl groups from RNA, while readers encompass binding proteins that can recognize specific methylation sites and facilitate the actions of both writers and erasers [4]. Currently, the body of research pertaining to m7G modification remains insufficient, and there exists no comprehensive report on m7G erasers. CBC and EIF4E are binding factors identified as m7G-caps readers, involving in various posttranscriptional processes of RNAs. CBC consists of the m7G-cap binding protein NCBP2 and the homologue NCBP1 [64]. Following m7G-caps modification by RNMT, mRNAs engage with NCBP2 and undergo a series of maturation steps, including splicing, CPA, and nuclear export [65]. Subsequently, cytoplasmic EIF4E assembles on the m7G-caps and recruits mRNAs to the translation machinery [66]. Notably, EIF4E is not limited to mRNAs; it can also engage in m7G-caps modification of certain ncRNAs, such as lncRNAs, miRNAs, and snoRNAs [67,68]. Among them, nuclear EIF4E can recognize m7G-caps of lncRNAs and promote their export, and m7G-capped pre-miRNAs are more likely to generate single 3p-miRNAs because of the biased selection by Ago. EIF4E is a prooncogenic molecule overexpressed and involved in many types of cancers. In AML, nuclear export of m7G-cap modification mRNA by EIF4E induce translation of c-MYC and Mcl-1, which are associated with relapse and poor prognosis [69]. Likewise, recent studies in osteosarcoma (OS) cells showed that EIF4E could enhance translation of cyclin D1 as well as mTOR pathway genes [70,71]. certain other family members of EIF4E can also exert critical roles in various cancer types through m7G capping, including the tumor suppressor EIF4E3 [72].
More recently, potential internal mRNA m7G-binding proteins have been identified via RNA pull-down assays. The results suggested that Quaking proteins (QKIs) could recognize and interact with m7G-modified oligonucleotides, then regulate stability and translation efficacy of mRNA under stress conditions. subsequently regulating the stability and translation efficiency of mRNA under stress conditions. In cancer cells, QKI7 has demonstrated the ability to mitigate chemoresistance to doxorubicin and sorafenib by reducing the translation of certain associated genes, including ROCK1, GSK3B, TEAD1, and IGF1R [52]. As the development of MeRIP-seq progresses, it is anticipated that more internal m7G readers will be identified, potentially shedding additional light on the mechanisms of m7G modification and its roles in cancer.
Other regulation factors in m7G modification
In addition to the determined m7G writers and readers discussed earlier, various internal and external factors play a pivotal role in influencing m7G processes and the expression of m7G regulators. These regulatory factors are commonly recognized as upstream mechanisms of m7G modification with significant implications in tumor initiation and progression. Internal factors involved in the regulation of m7G modification encompass oncogenic molecules or pathways, along with epigenetic modifications. Prior investigations have suggested that the classical m7G writer METTL1 may undergo transcriptional upregulation by the transcription factor (TF) SP1, thereby stabilizes CDK14 mRNA and contributes to the progression of prostate cancers [53]. Whole exon sequencing in nasopharyngeal cancer (NPC) has elucidated the downregulation of METTL1 by the pivotal tumor suppressor ARNT [73]. Similarly, the transcription of WDR4 was noted to be activated by c-MYC, thereby potentially fostering HCC progression [74]. Furthermore, the analyses of m7G genetic variation revealed that the cope number variation (CNV) and single nucleotide variation (SNV) of m7G regulators were prevalent in various cancers, and researchers also demonstrated the correlation between the methylation level and expression of m7G regulators [75]. Regarding the external factors, m7G modification processes can be triggered under stress conditions, such as ionizing radiation (IR), heat shock and drug treatments. Recent studies have indicated that chemotherapeutic treatments in diverse tumor cells can induce mRNA m7G modification mediated by METTL1 and QKI7. Furthermore, radiotherapy in HCC has been associated with the overexpression of METTL1[52,76]. Both internal and external regulatory factors predominantly influence m7G regulators, and these investigations hold promise for devising effective strategies for combination therapies across various cancer types.
The roles of m7G modification in immune cell biology
Immune system can exert its function of immune surveillance and immune response against tumor cells, which play crucial roles in proliferation, metastasis and treatment resistance of cancers [77,78]. Tumor immune microenvironment (TIME) consists of tumor, stromal and immune cells including adaptive and innate immune cells [79]. Current studies have showed remarkable enrichment of m7G regulators in biological processes in TIME, especially immune infiltration [6]. Here, we reviewed the studies of prognostic models based on m7G levels and concluded activation or accumulation of immune cells mediated by m7G modification. The detailed information is also showed in Table 1.
Biological functions and mechanisms of m7G modification in tumor-infiltrating immune cells
| Immune cells | Cancer types | Related m7G regulators | Functions and mechanisms | Ref. | |
|---|---|---|---|---|---|
| T cells | CD4+ cells | HCC | METTL1 | More infiltration of Tregs and activated memory T cells by METTL1 overexpression | [97] |
| LUAD | / | More infiltration of Tregs and Th cells in low-risk patients | [104] | ||
| Breast cancers | METTL1 | More infiltration of Tregs by METTL1 overexpression | [97] | ||
| / | More infiltration of Tregs and Th cells in high-risk patients | [132] | |||
| GC | / | More infiltration of Th cells in low-risk patients | [88] | ||
| HNSCC | METTL1 | Decreased infiltration of Tregs in METTL1-KO mice | [99] | ||
| CD8+ cells | LUAD | WBSCR22 | More infiltration of activated CD8+ cells | [101] | |
| GC, prostate cancers, STS | / | More infiltration of CD8+ cells in low-risk patients | [86,88,89] | ||
| HNSCC | EIF4E3, NCBP2 | More infiltration of CD8+ cells in low-risk patients | [85] | ||
| B cells | Naïve B cells | CRC, LUAD | / | More infiltration of naïve B cells in low-risk patients | [104,105] |
| Cutaneous melanoma | / | More infiltration of naïve B cells in high-risk patients | [106] | ||
| Memory B cells | GC, HCC, bladder cancer | / | More infiltration of memory B cells in low-risk patients | [107-109] | |
| Plasma cells | GC, HCC | / | More infiltration of plasma cells in low-risk patients | [108,109] | |
| MDSCs | ICC | METTL1 | More CXCL8 translation and consequent PMN-MDSC infiltration regulated by METTL1 | [112] | |
| HCC | METTL1 | Activated TGF-β pathway and more PMN-MDSC infiltration regulated by METTL1 | [76] | ||
| NUDT16 | More infiltration of MDSCs in high-risk patients | [114,115] | |||
| SKM | EIF4E3, IFIT5 | More infiltration of MDSCs in low-risk patients | [116] | ||
| Macrophages | Prostate cancer | METTL1 | Suppressed IFN signaling pathway and M2 macrophages polarization mediated by METTL1 | [123] | |
| LUAD | WDR4 | Enhanced expression of CD73 and infiltration of CD206+ M2 macrophages | [98] | ||
| CRC, ovarian cancer | / | More infiltration of M1 macrophages in low-risk patients | [96,126] | ||
| Bladder cancer | / | More infiltration of M1 macrophages in high-risk patients | [127] | ||
| GC, PDAC | / | More infiltration of M2 macrophages in high-risk patients | [109,125] | ||
| Monocytes | CRC, bladder cancer | / | More infiltration of monocytes in low-risk patients | [127,128] | |
| Neutrophils | PDAC | FN1, ITGB1 | More infiltration of neutrophils in high-risk patients | [125] | |
| DC | HCC | NUDT16 | More infiltration of DCs in high-risk patients | [115] | |
| CRC, LUAD | / | More infiltration of DCs in low-risk patients | [128,133] | ||
| NK cells | CRC, SKM, LUAD | / | More infiltration of NK cells in low-risk patients | [116,128,138] | |
M7G modification and its regulators exert important functions in immune cell biology of TIME. The infiltration of T cells, B cells, MDSCs, macrophages, monocytes, DCs and NK cells has been reported to be influenced by m7G modification, and several different types of cancers are involved.
T cells
T cells constitute pivotal components of the adaptive immune system, encompassing distinct subtypes such as CD4+ and CD8+αβ T cells, γδ T cells, and natural killer T (NKT) cells [80]. Upon recognition of cognate antigens by T cell receptors and subsequent stimulation by cytokines, naïve CD4+ and CD8+ T cells undergo a series of intricate biological processes. These processes encompass maturation, differentiation into effector or memory cells, activation, and subsequent migration to specific anatomical locations [81,82]. Prior studies have extensively explored various T cell subsets in the TIME. CD8+ T cells, recognized as highly responsive and effective cytotoxic T lymphocytes (CTLs), exert their antitumor effects by targeting tumor cells, predominantly through the secretion of cytotoxic proteins and the induction of cellular apoptosis [83,84]. In general, the infiltration of CD8+ cells has been reported to display a negative correlation with m7G levels and is downregulated in high-risk patients. In the context of head and neck squamous cell carcinoma (HNSCC), cluster analysis utilizing the ESTIMATE and CIBERSORT algorithms has revealed a higher proportion of CD8+ T cells in the low-risk group [85]. Studies conducted in CRC, prostate cancer, soft tissue sarcoma (STS), gastric cancer (GC), and ovarian cancer have suggested a similar regulatory pattern [86-90]. However, CD4+ cells displayed varying abundances across different cancer subtypes and types. Helper T (Th) cells originate from activated CD4+ T cells and differentiate through distinct surface markers such as Interferon (IFN)-γ, GATA-3, and various interleukins [91-93]. Among them, Th1, Th2, and Th17 cells appeared to be more stimulated in low-risk patients with GC and prostate cancer, while the analysis in STS patients indicated a reduced recruitment of Th cells [86,88,89]. Regulatory T cells (Tregs) represent a subset of CD4+ T cells with distinctive significance in TIME. Renowned for their suppressive capabilities, Tregs exert inhibitory effects on effector T cells, thereby contributing to immune tolerance towards tumor cells [94,95]. Clustering analysis performed in the context of CRC and ovarian cancers have revealed a heightened prevalence of Tregs within the high-risk patient cohort. This elevated accumulation of Tregs aligns with an associated decline in overall survival [90,96].
Numerous studies in past decades reported that m7G regulators could influence the infiltration, maturation and activation of T cells in TIME. Among these regulators, the METTL1 and WDR4 are prominently studied m7G writers within the context of T cell biology. In a comprehensive analysis of pan-cancers, overexpression of METTL1 and WDR4 was correlated with more Tregs infiltration in diverse types of cancers [97]. Mechanistically, WDR4 was reported to elevate intertumoral Tregs and foster a suppressive TIME by promoting the degradation of the pleiotropic tumor suppressor promyelocytic leukemia protein and inducing CD73 expression in lung adenocarcinoma (LUAD) [98]. Moreover, METTL1 has been implicated in augmenting the proportion of Tregs in HNSCC through promoting PI3K protein translation and AKT/mTOR signaling pathway activation in m7G-dependent manner [99]. Additionally, the m7G-caps writer RNMT was reported to regulate CD4+ T cells activation. RNMT catalyzes the m7G-caps formation in genes and snoRNAs required for ribosome biogenesis, thus enhancing the translation process in activated T cells [100]. WBSCR22 could also participate in activation of CD8+ T cells in LUAD, and the m7G-related cap-binding genes NCBP2 and EIF4E3 were suggested to involve in infiltration of CD8+ T cells in HNSCC [85,101]. These findings indicated the role of m7G modification and its regulators in T cells biology of TIME.
B cells
B cells perform crucial roles in adaptive immunity, encompassing antibody production, antigen recognition, and cytotoxicity [102]. Tumor-infiltrating B cells can be categorized into distinct subtypes based on their stages of differentiation, including naïve B cells, activated B cells, memory B cells, and plasma cells [103]. The extent of immune infiltration by these subtypes varies across different cancer types. For example, a negative correlation was observed between the infiltration of naïve B cells and the risk score for an unfavorable prognosis in CRC and LUAD. However, clustering analysis in cutaneous melanoma indicated a higher level of naïve B cell infiltration in the high-risk group [104-106].
Upon stimulation by antigens, B cells undergo activation and subsequently differentiate into two distinct cell types: memory B cells and plasma cells. Memory B cells accumulation was negatively correlated with m7G levels in GC, HCC and bladder cancer, while LUAD patients appeared opposite results [104,107-109]. Plasma cells are effector cells differentiated from activated B cells which produce antibodies. Generally, tumor-infiltrating plasma cells are abundant in low-risk groups, which are also associated with better prognosis [109].
Myeloid-derived suppressor cells (MDSCs)
MDSCs represent a population of immature myeloid cells with the capacity to suppress T-cell activity, ultimately contributing to the establishment of an immunosuppressive TIME in the advanced stages of cancer [110]. MDSCs are characterized by co-expression of CD33, CD34, CD11b and IL-4Rα, and identified as two major subsets: polymorphonuclear- (PMN-) and monocytic- (M-) MDSCs [111]. The classical m7G writer METTL1 was reported to regulate the accumulation of PMN-MDSCs in various cancers. In ICC, it was demonstrated that abundance of PMN-MDSCs with CD15 expression was increased in advanced tumors. Mechanistically, researchers revealed that METTL1 could promote CXCL8 translation by tRNA m7G modification, thereby activate CXCL8/CXCR2 chemokine pathways and induce the recruitment of PMN-MDSCs in TIME [112]. Studies in HCC reported similar function of METTL1 in regulating PMN-MDSCs infiltration. The translation of TGF-β2 was upregulated by METTL1-dependent tRNA m7G modification, which could induce the differentiation of peripheral blood mononuclear cells (PBMCs) into PMN-MDSCs and increase their infiltration, thereby suppress the function of CD8+ T cells and promote HCC progression [76,113]. Moreover, clustering analysis in HCC demonstrated that the m7G modification level positively correlated with abundance of MDSCs in TIME, and some novel regulators such as NUDT16 and KIF2C were involved in MDSCs regulation [114,115]. On the contrary, comprehensive analysis based on m7G-related genes in skin cutaneous melanoma (SKM) suggested the higher overall survival regulated by EIF4E3 and IFIT5, which was associated with more immune infiltration including MDSCs [116].
Macrophages
Macrophages, derived from monocytes, are integral innate immune cells [117]. They hold a pivotal role within the TIME and serve as pivotal regulators across diverse biological processes including epithelial to mesenchymal transition (EMT), angiogenesis, ECM remodeling, and instigation of immunosuppressive responses [118,119]. Tumor-associated macrophages (TAM) possess two polarization settings: pro-inflammatory M1 macrophages and anti-inflammatory M2 macrophages [120]. Being the predominant macrophages infiltrating tumor tissues, M2 macrophages contribute to the establishment of a suppressive TIME by secreting inhibitory cytokines, inducing apoptosis in immune cells, directly attenuating TCR and BCR signaling through PD-1/PD-L1 axis, and collaborating with other immune cells to exert suppressive functions [121,122]. The classical m7G writers METTL1 and WDR4 have been implicated in the polarization of macrophages towards the M2 phenotype and the establishment of a suppressive TIME. In prostate cancers, researchers found that METTL1-mediated tRNA m7G modification could impede the biogenesis of tRNA fragment, consequently diminishing the translation of transcripts associated with IFN signaling pathway and promote macrophage polarization towards the M2 phenotype [123]. Likewise, overexpression WDR4 could contribute to enhanced infiltration of CD206+ M2 macrophages, as well as reduced CD8+ T cells recruitment, which induced a suppressive TIME in LUAD [98]. Moreover, the clustering analysis showed a positive correlation between m7G modification level and M2 macrophages infiltration across various cancers [106,109,124,125]. M1 polarization of macrophages were usually conventionally with favorable outcomes. The prognostic models of ovarian cancers, CRC, HCC and HNSCC based on m7G regulators indicated more recruitment of M1 macrophages in low-risk group of patients [85,96,115,126]. However, contradictory outcomes were observed in bladder cancers [107,127]. In addition, as precursor cells of macrophages, monocytes infiltration was reported to be negative correlated with m7G levels in CRC, bladder cancers and prostate cancers [89,127,128]. These results are consistent with the role of monocytes in mononuclear phagocyte system.
Neutrophils
Neutrophils serve as the primary defense mechanism during inflammatory conditions and act as effector cells to combat pathogens [129]. In various types of cancers, neutrophils show the double-edged roles which can either inhibit tumor progression through cytotoxic activity or promote immunosuppression in cancers [130,131]. Likewise, the influence of neutrophils infiltration by m7G related genes is still controversial. In pancreatic ductal adenocarcinoma (PDAC), researchers suggested that m7G scores based on expression of m7G regulators were positively correlated with neutrophils infiltration, and patients with high m7G scores and increased number of neutrophils appeared higher malignancy degrees and worse prognosis. PPI network showed that FN1 and ITGB1 were the core genes in regulating m7G [125]. Similar mechanisms were demonstrated in breast cancers and bladder cancers [127,132]. However, it was reported in LUAD and cutaneous melanoma that neutrophils were less infiltrated in group of high m7G levels [56,133]. Except for neutrophils, further studies are needed in eosinophils and basophils, which are members of granulocytes as well.
Others
Nature killer (NK) cells, dendritic cells (DCs) and mast cells belong to innate immune cells which participate in TIME formation. Among them, NK cells serve as the primary defense against cancer, effectively targeting tumor cells without the need for prior sensitization [134]. Elevated NK cell abundance is observed among low-risk patients exhibiting favorable prognostic outcomes, typically linked to reduced m7G modifications [115,116,128]. DCs function as antigen-presenting cells (APCs), playing a pivotal role in identifying tumor cells and presenting antigens to naïve T cells [135,136]. In HCC, patients cluster with m7G levels upregulation suggested immunosuppressive phenotypes and poor prognosis. Meanwhile, these patients showed increased infiltration of DCs as well as decreased activated T cells [115]. However, CRC, LUAD and cutaneous melanoma showed opposite results that m7G modification in high-risk patients could suppress DCs infiltrating [56,128,133]. These findings align with the double-edged functions of DCs in TIME. Furthermore, mast cells can also accumulate into TIME and exert both pro- and antitumorigenic functions [137]. A risk model based on m7G regulated genes in LUAD suggested the connection of advanced stages tumor and mast cells infiltration, and studies indicated different recruitment in different types of cancers [107,108,138].
M7G regulate TIME by affecting tumor and stromal cells
Except for various types of immune cells, tumor cells and stromal cells are also important components of TIME [139]. Investigating the intricacies of immune cell biology governed by m7G modification, scholars have dedicated comparable attention to scrutinizing the impact of m7G within tumor cells and stromal cells. Recent research indicates that m7G modification exerts its influence on both the tumor and stromal constituents of TIME through distinct mechanisms. These mechanisms encompass alterations in the expression of immune checkpoints, secretions of related bioactive factors, as well as the function and communication of stromal cells. A detailed exposition of these aspects is provided below.
M7G modification impact the immune checkpoints of tumor cells
Immune checkpoints are cell surface molecules of T cells, APCs and tumor cells, which play pivotal roles in modulating immune response [140,141]. Based on their diverse functions in T cell activation, immune checkpoints can be categorized into co-stimulatory molecules such as CD28, CD80/CD86, and co-inhibitory ones like PD-1/PD-L1 and CTLA-4[142]. The co-inhibitory immune checkpoints function to diminish the immunogenicity of tumor cells, facilitating their evasion from immune surveillance, which are regarded as crucial immunotherapy targets [143]. Specifically, PD-1 and CTLA-4 are predominantly located on the surface of activated T cells, while PD-L1 is expressed on tumor cells in TIME [144]. Previous studies of risk models across various cancers indicated that m7G regulators could impact the expression of several immune checkpoints, particularly within tumor cells. In a prognostic risk model of LUAD based on m7G regulators, it suggested that PD-L1 expression was obviously upregulated in high-risk group of patients. Meanwhile, PD-1 and CTLA-4 were overexpressed as well [133,138]. Analogous studies in GC, CRC and lower-grade glioma (LGG) showed the overexpression of PD-L1 in patients with high m7G level [105,145,146], and the other immune checkpoint of tumor cells, CD276 showed same outcomes [147]. Consequently, m7G modification emerged as a predictive factor for the immunological response to anti-PD-L1 therapy [116,146]. These findings underscored the influence of m7G modification on the expression of immune checkpoints in tumor cells, positioning it as a potential biomarker or therapeutic target for immunotherapy.
M7G modification regulate the secretion of immune related bioactive factors
Except for immune checkpoints located on the surfaces of tumor and immune cells, paracrine signaling through secretion of cytokines, chemokines, IFNs or other immune related bioactive factors is critical for immune response and formation of TIME [148]. In recent years, an increasing number of researches have focused on the role of m7G modification in regulating these factors and the downstream signaling pathways. The m7G writer METTL1 was reported to participate in various processes. For example, polyribosome mRNA-seq in ICC cells showed that knockout of METTL1 could reduce translation ratio of the chemokine CXCL8 by affecting the m7G-decoded codon frequency, thereby inhibit the migration of MDSCs to tumors [112]. Likewise, researchers also verified the translational activation of TGF-β2 mediated by METTL1-related tRNA m7G modification in pathway analysis of HCC cells [76]. And the secretion of TGF-β2 could lead to accumulation of PMN-MDSCs which shaped suppressive TIME and fostered tumor immune evasion [149]. Moreover, several pro- and anti-inflammatory cytokines were involved in m7G modification as well. Cytokine composition analysis in prostate cancer cells indicated that the loss of METTL1 could facilitate the secretion of IFN, tumor necrosis factor α (TNF-α) and granulocyte-macrophage colony-stimulating factor (GM-CSF), while downregulate release of macrophage colony stimulating factor (M-CSF), IL10 and IL13[123]. In conclusion, m7G modification played crucial roles in translation and secretion of numerous immune related factors, as well as the regulation of downstream signaling pathways. These processes could also influence the intercellular communication in TIME.
M7G modification influence the function and communication of stromal cells
Stromal cells in TIME consist of blood and lymphatic endothelial cells, mesenchymal stem cells (MSCs), cancer-associated fibroblasts (CAFs), pericytes and neurons [150]. These entities, together with the extracellular matrix (ECM) make up the stromal component. Analogous to immune cells, the function and phenotype of stromal cells have been reported to be influenced by m7G modification and related regulators. In the risk models based on m7G-related genes, researchers utilized the ESTIMATE algorithm to assess stromal scores, thereby evaluating the function of stromal cells in TIME. The results showed higher stromal scores in high-risk subgroups of patients with HCC, GC and ESCC, while SKM and LUAD patients represented opposite outcomes [104,116,145,151,152]. These findings indicated diverse activation and accumulation of stromal cells in diverse cancers. Moreover, single cells RNA-seq analysis in HNSCC also revealed that METTL1 knockout could impair direct communication between stromal cells and epithelial cells via inhibition of IL-1b and its receptors [99]. Specific to each subtype, the infiltration of CAFs was demonstrated to be positively correlated with the m7G regulator KIF2C, and eIF4E could promote the oncogenic transformation of fibroblasts by facilitating nuclear export of cyclin D1 mRNAs [71,114]. Previous studies revealed that the endothelial cell line HUVEC could be shaped by METTL1. Mechanistically, METTL1 could induce the m7G modification of VEGFA mRNA, thus promotes HUVECs migration and angiogenesis [153]. However, further investigations are warranted to explore the interaction between m7G modification and other subtypes of stromal cells.
M7G modification influence the efficacy of adjuvant therapy
Surgical resection remains the primary therapeutic approach for the majority of solid tumors among the various treatment modalities available for cancer management. In recent years, significant advancements in our understanding of tumor biology have paved the way for the integration of adjuvant therapies, including radiotherapy, chemotherapy, immunotherapy, and combination therapy. These adjunctive treatments aim to reduce tumor burden, downgrade primary tumor staging, and increase the feasibility of surgical intervention [154]. However, the effectiveness of adjuvant therapies remains constrained by factors such as drug susceptibility, tolerance, and the potential for tumor recurrence. Building upon prior research, it has become evident that m7G modification plays a pivotal role in modulating the outcomes of various adjuvant therapies across different cancer types. These findings are comprehensively discussed in the following sections and summarized in Table 2.
The roles of m7G modification in adjuvant therapies of cancers
| Therapeutic methods | Cancer types | Related m7G regulators | Functions and mechanisms | Ref. | |
|---|---|---|---|---|---|
| ICB therapy | Anti-PD-L1, anti-PD-1, anti-CTLA-4 | HCC, PDAC, SKM, GC, LGG, etc. | / | Increased treatment responses in patients of low-risk groups | [109,115,116,125] |
| Two-drugs combination therapy | PDAC | / | Better efficacy in patients of high-risk groups | [125] | |
| Anti-PD-1 | ICC | METTL1 | Enhanced efficacy by co-blockade of METTL1 and downstream chemokine pathways | [112] | |
| Anti-PD-1, anti-CTLA-4 | Prostate cancer | METTL1 | Enhanced efficacy by METTL1 inhibition | [123] | |
| Chemotherapy | Cisplatin and docetaxel | NPC | METTL1/WDR4 | Promoted chemoresistance by METTL1 and WNT/β-catenin pathway | [73] |
| Doxorubicin | OS | METTL1/WDR4 | Promoted chemoresistance by METTL1 | [171] | |
| HCC, cervical cancer | QKI7 | Promoted chemosensitivity by QKI7-mediated inhibition of Hippo pathways | [52] | ||
| Anti-BRAF and anti-MEK compounds | Melanoma, CRC, thyroid cancer | EIF4E | Decreased resistance by combination therapy with inhibition of EIF4E-EIF4G interaction | [173] | |
| Ribavirin | AML | EIF4E | Clinical benefits of m7G-cap mimic Ribavirin by targeting EIF4E | [172] | |
| Lenvatinib | HCC | WDR4 | Promoted chemoresistance by WDR4 and TRIM28 | [174] | |
| HCC | METTL1 | Promoted chemoresistance by METTL1 and EGFR pathway | [175] | ||
| Sorafenib | HCC | WDR4 | Promoted chemoresistance by MYC-targeted WDR4 and CCNB1 | [74] | |
| Radiotherapy | HCC | METTL1 | Promoted IR resistance by METTL1-mediated NHEJ-based DNA repair | [176] | |
| RFA | HCC | METTL1 | Immunosuppressive TME and residual HCC progression by upregulation of METTL1 in post-RFA recurrent tumor | [76] | |
| HCC | METTL1 | Increased post-RFA HCC metastasis by METTL1-mediated upregulation of SNAI1/SLUG axis | [177] | ||
m7G modification plays a pivotal role in modulating the effectiveness of various adjuvant therapies, including ICB therapy, chemotherapy, radiotherapy and RFA. The different cancer types and concrete mechanisms are shown above.
Immune checkpoint blockade (ICB) therapy
As ICB therapy has demonstrated substantial efficacy and promise in cancer treatment over the past several decades, immune checkpoint inhibitors (ICIs) have found application in clinical practice for attenuating the inhibitory signals hindering T-cell activation and promoting anti-tumor immune responses [155]. In contemporary clinical practice, ICIs predominantly encompass agents targeting CTLA-4, PD-1, and PD-L1. The pioneering success in cancer immunotherapy was achieved with the anti-CTLA-4 antibody, Ipilimumab. Rigorous double-blind phase II trials conducted in advanced melanoma demonstrated that Ipilimumab monotherapy significantly prolonged overall survival and exhibited a dose-dependent effect on antitumor efficacy [156]. Recent investigations have uncovered the therapeutic potential of Ipilimumab not only in monotherapy but also in combination with other ICIs or chemotherapeutics [157-159]. Anti-PD-1 antibodies are ICIs with most variety of approval oncology therapeutic products in clinic application, such as Pembrolizumab and Nivolumab [160]. Both monotherapy and combination based on these inhibitors showed prognostic benefit in patients of various cancers, and the treatment effect was enhanced with overexpression of PD-L1[161-163]. In order to improve the efficacy and safety of ICB therapy, researchers nowadays have paid more attention to bispecific antibodies which simultaneously target 2 different immune checkpoints, and there have been numerous associated phase I or phase II clinical trials in types of cancers [164-166]. Apart from PD-L1 expression, tumor mutation burden (TMB) and tumor immune dysfunction and exclusion (TIDE) were used to assess the treatment response in the prevailing views. TMB represent the neoantigens generated by the mutations in tumor, which can induce T cells recognition and immune response [167]. TIDE reflect the induction of T cells dysfunction and prevention of CTLs infiltration which lead to immune evasion in TIME [168]. Generally, higher TMB and lower TIDE may predict enhanced efficacy of ICB therapy. Previous evidence suggested that m7G modification and its regulators could influence the response to ICB therapy in many cancers. On the one hand, m7G scores based on m7G levels and prognostic risks were important biomarker in ICB therapy. Clustering analyses conducted in GC, CRC, LUAD and bladder cancers revealed a noteworthy association between higher m7G risk scores and lower TMB. This correlation was further linked to diminished responses to ICB therapy and a poorer prognosis for patients. Additionally, TIDE scores demonstrated a consistent negative correlation with m7G levels across various cancer types [105,109,138,169]. These observations underscored the potential of m7G as a valuable biomarker for predicting responses to ICB treatment. For the concrete ICB therapy strategies, researchers examined the predictive value of m7G signature in different immunotherapy cohorts. According to the anti-PD-1 and anti-PD-L1 immunotherapy cohorts, low-risk group of patients in HCC, SKM, LGG and bladder cancers showed significant clinical benefits for anti-PD-1 or anti-PD-L1 treatment [115,116,146,170]. However, the scenario is nuanced when considering combination therapy. In PDAC, the efficacy of anti-PD1 and anti-CTLA-4 combined treatment in high-risk group patients was better than low-risk group, while single ICI showed opposite outcomes [125]. These findings underscored that ICB therapy efficacy was negative correlated with risk scores based on m7G levels, while the regulation in combination therapy was still complicated.
On the other hand, some specific m7G-related genes, especially m7G writers play important roles in ICB therapy. METTL1 stands as the most extensively studied m7G methyltransferase within the context of tumor immunity. In ICC, researchers constructed hydrodynamic transfection models of METTL1 knockout mice. The results suggested that METTL1 knockout could increase the proliferation and antitumor activity of T cells, as well as boost the efficacy of anti-PD-1[112]. More recently, investigations in prostate cancer indicated that knockout of METTL1 could enhance the sensitivity of ICB therapies containing anti-PD1 and anti-CTLA-4 antibodies [123]. Furthermore, a comprehensive analysis of pan-cancers also explored whether METTL1 could influence the efficacy of anti-PD-L1 therapy. The expression of PD-L1 was in negative proportion to METTL1, but patients with higher METTL1 expression showed more responses to anti-PD-L1 therapy. These findings revealed that METTL1 may not be suitable target of combination therapy with anti-PD-L1[97]. In conclusion, m7G modification as well as the related regulators are important biomarkers and potential targets in ICB therapy, and investigation of METTL1 inhibitors may provide innovative strategies to enhance the efficacy of ICIs like anti-PD1 and anti-CTLA-4.
Chemotherapy
Chemotherapy is a cornerstone treatment modality across a wide spectrum of cancer types, and resistance to chemotherapy stands as the foremost determinant of unfavorable prognoses. Notably, M7G modification has emerged as a factor with significant influence on the resistance of traditional chemotherapy in various cancers. For example, in NPC, m7G modification of tRNAs catalyzed by METTL1/WDR4 complex could accelerate codon recognition and mRNA translation, which contributed to activation of WNT/β-catenin pathway and subsequently promoted chemoresistance of NPC cells to cisplatin and docetaxel [73]. Similarly, m7G tRNA modification in OS heightens chemoresistance to doxorubicin by increasing mRNA translation [171]. Except for m7G writers, some m7G readers including EIF4E and QKIs have also been implicated in the regulation of chemoresistance. The classical antiviral drug Ribavirin is a mimic of m7G-caps, which can inhibit nuclear export of EIF4E. Researchers applied Ribavirin in M4/M5 AML patients and showed its clinical benefits [172]. Boussemart et al. uncovered that EIF4E cap-binding protein could form a complex with EIF4G and EIF4A, thereby suppressing the therapeutic efficacy of anti-BRAF and anti-MEK compounds in melanoma, colon, and thyroid cancer cell lines. Subsequent investigations unveiled that the inhibition of EIF4E-EIF4G interaction and translation activation could overcome the chemoresistance of these chemotherapy drugs [173]. Moreover, QKI7-mediated internal mRNA m7G modification were involved in chemoresistance upon doxorubicin. The overexpression of QKI7 in HeLa and HCC cells could enhance doxorubicin-induced apoptosis and sensitized cancer cells to chemotherapy drugs [52]. These findings underscored diverse functions of m7G regulators in tumor initiation and progression as well.
Apart from the traditional chemotherapy drugs, previous studies indicated that sensibility and resistance of molecular targeted therapies could also be regulated by m7G modification. For instance, the tyrosine kinase inhibitor (TKI) Lenvatinib is the first-line therapy for HCC, and the Lenvatinib-resistant patients showed increased level of m7G modification [174]. Further studies demonstrated that the resistance of Lenvatinib were enhanced by translational activation of EGFR, which was dependent on tRNA m7G modification catalyzed by METTL1/WDR4 complex [175]. Likewise, the other TKI in HCC therapy, Sorafenib was reported to be associated with WDR4. WDR4 induced translation of CCNB1 mRNA by tRNA m7G modification, thus promoted sorafenib resistance [74]. Furthermore, it is imperative to undertake further investigations to unravel the intricate interplay between targeted therapies and m7G modification in various other cancer types.
Radiotherapy and radiofrequency ablation (RFA)
Radiotherapy and RFA belong to local treatments of cancers. Notably, radiotherapy has gained prominence as an essential therapeutic approach in the management of HCC. The efficacy of radiotherapy is based on DNA damage, thus the radioresistance of cancer cells are associated with DNA DSB repair. Liao et al. demonstrated that METTL1 could render HCC cells resistant to IR by upregulating DNA-PKcs and DNA ligase IV translation and promoting non-homologous end-joining (NHEJ) mediated repair [176]. RFA is an effective local therapeutic strategy of various cancers, and the post-RFA tumor recurrence is the dominant factor to influence the therapeutic effects. Investigation of RFA and m7G modification mainly concentrated on HCC. Researchers have demonstrated that insufficient RFA could upregulate METTL1 and lead to immunosuppressive TME, which resulted in PMN- MDSCs and residual HCC progression [76]. Furthermore, further studies by the same team indicated that METTL1-mediated tRNA m7G modification could facilitate post-RFA HCC metastasis by inducing translation of SNAI1 and SLUG, which provide potential therapy target to decrease recurrence rate [177]. To achieve a comprehensive understanding of m7G modification in conjunction with RFA, it is imperative to conduct further research encompassing various cancer types in the future.
Conclusion and prospects
In this comprehensive review, we mainly focused on the m7G modification and its pivotal role in the regulation of the TIME and adjuvant therapies. The m7G modification, a prevalent RNA modification in eukaryotes, has garnered substantial attention from researchers in recent decades. This heightened interest can be attributed to the advent of advanced methodologies for the detection of m7G, including LC-MS/MS, m7G-seq, m7G-MeRIP-seq and m7G-miCLIP-seq. These innovative approaches have shed light on the pervasive nature of m7G modification, implicating its involvement not only in mRNAs but also in numerous ncRNAs, including tRNAs and rRNAs. To date, clearly reported regulators of m7G modification contain the m7G writer complexes such as RNMT/RAM, METTL1/WDR4 and WBSCR22/TRMT122, the m7G-caps readers CBC and EIF4E, as well as internal m7G binding protein QKI. Despite some m7G-related genes and bridge genes were reported in many integrated analyses and prognostic models, their specific mechanisms of regulating m7G modification were elusive without concrete supporting evidence. Apart from these m7G regulators, some other internal and external regulation factors including aberrant epigenetic modification, drug or IR treatments, as well as some oncogenic molecules and signaling pathways are also involved in tumor initiation and progression.
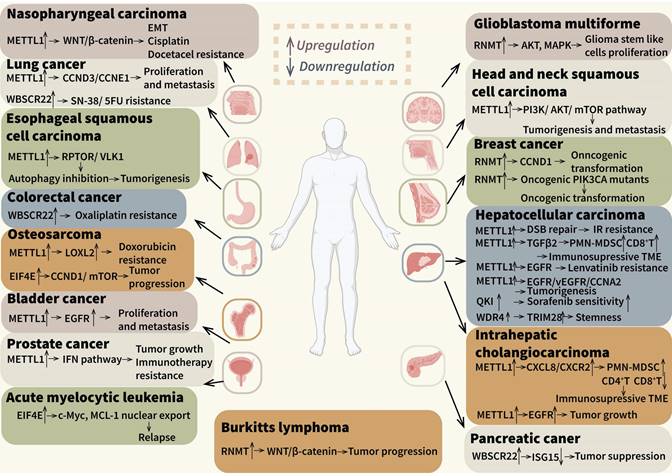
We have systematically reviewed the roles of m7G modification in regulating tumor initiation and progression of different malignancies, including various solid tumors, leukemia and lymphoma, as it illustrated in Figure 4. (Figure 4). Firstly, distinct cancer-associated m7G mechanisms manifest in different cancer types. Dysregulation of m7G-caps modification emerges as a primary mechanism associated with tumor progression and relapse of breast cancers and hematological malignancies, while aberrant internal m7G is more prevalent in HCC and ICC [48,172,178]. Secondly, diverse downstream pathways like EGFR/MAPK, AKT/mTOR and WNT/β-catenin were involved in different cancer types, which might cause different pathological behaviors [73,99,175]. Furthermore, we observed varied effects on tumor treatment outcomes. It was disclosed that m7G modification could mediate IR resistance and relapse after RFA, but researches in other tumors mainly focused on chemotherapy like platinum drugs. These findings were also reported in previously published reviews. Notably, the regulation of immune cell biology within the TIME was implicated. Herein, we concentrated on the risk models based on m7G-related genes or different expression genes and some mechanistic researches. The infiltration and recruitment of immune cells including T cells, B cells, MDSCs, macrophages and neutrophils were correlated with m7G modification levels and regulators, which simultaneously showed different outcomes in distinct types of cancers. In addition, the stromal cells and tumor cells in TIME can also be regulated by m7G modification, which can influence immune checkpoints expression, cytokines and chemokines secretion, as well as the functions of stromal cells. Furthermore, researchers have suggested that m7G modification is associated with the efficacy of many adjuvant therapies. In ICB therapies, m7G-related risk scores were negative correlated to curative activity of ICIs, and the m7G writer METTL1 could inhibit efficacy of ICB therapies. Nevertheless, challenges persist in terms of efficacy and safety in the monotherapy of ICIs, necessitating the exploration of combination therapies in various cancers [179]. Mechanistically, chemotherapy drugs and radiation can activate immunostimulatory signals via eliminating the tumor cells, thereby promoting the cytotoxicity of immune effector cells and suppressing the formation of suppressive TIME [180]. These observations underscored the imperative for further investigations into alternative anticancer treatments. Previous studies demonstrated that chemotherapy, radiotherapy and RFA could also be influenced by m7G regulators METTL1, EIF4E and QKI7. Apart from the solid tumors, some hematologic tumors like AML also involved in m7G-related combination therapies.
The role of m7G modification and its regulators in diverse cancers. M7G modification and its regulators have been reported in many types of cancers. The related cancers and downstream genes and pathways are showed in the figure.
Previous investigations have demonstrated the significance of m7G and its regulators as important biomarkers for diagnosis, prognosis and immunotherapy in various cancers. Overall, m7G levels are positively correlated with the TMN stages in most cancers, and high m7G scores in patients usually predict worse prognosis. Moreover, m7G levels may affect the TMB and TIDE scores, as well as the expression of immune checkpoints, thereby influence the efficacy of immunotherapy [75]. In terms of m7G regulators, METTL1 and WDR4 have been regarded as pivotal oncogenic molecules associated with poor overall studies, which also provide possibilities for early diagnosis and treatment [14,51]. Simultaneously, other regulators and m7G-related genes have showed the expression and functions in various cancers, with only a few members showing downregulation in tumor tissues and association with clinical benefit. These findings have also revealed the oncogenic role of m7G in most of cancers. However, relevant clinical trials targeting m7G and m7G regulators are still lacking, and most of researches to date are based on mechanistic exploration and clustering analysis. From another perspective, the research status indicates great potential for targeting m7G modification in clinic application, especially combination therapies. For example, knocking down METTL1 by using inhibitors, antibodies or nanomedicine methods can be served as promising therapy strategies, as mechanistic studies have illustrated that the efficacy of ICIs and certain chemotherapeutic agents is augmented in METTL1 knockout mice. This suggests a synergistic effect when combining these therapeutics with m7G-targeted therapy. As targeted therapy continues to advance, novel inhibitors of m7G regulators may find application in adjuvant therapy alongside ICIs and other targeted drugs. In summary, m7G modification emerges as a pivotal factor in regulating the TIME and enhancing adjuvant therapy efficacy. Consequently, it holds promise as both a potential biomarker and a target for combination therapy.
Abbreviations
M7G: N7-methylguanosine; m6A: N6-methyladenosine; mRNA: messenger RNA; tRNA: transfer RNA; rRNA: ribosomal RNA; ncRNA: non-coding RNA; miRNA: microRNA; snoRNA: small nucleolar RNA; RNMT: RNA guanine-7 methyltransferase; RAM: RNMT-activating miniprotein; METTL1: methyltransferase-like 1; WDR4: WD repeat domain 4; WBSCR22: Williams-Beuren syndrome chromosomal region 22; TRMT112: tRNA methyltransferase activator subunit 11-2; CBC: cap-binding complex; EIF4E: eukaryotic translation initiation factor 4E; TIME: Tumor immune microenvironment; MDSC: myeloid-derived suppressor cell; TAM: tumor-associated macrophages; DC: dendritic cell; ICB: immune checkpoint blockade; ICI: immune checkpoint inhibitors; TKI: tyrosine kinase inhibitor; HCC: hepatocellular carcinoma; ICC: intrahepatic cholangiocarcinoma; GC: gastric cancer; CRC: colorectal cancer; LUAD: lung adenocarcinoma; PDAC: pancreatic ductal adenocarcinoma; HNSCC: head and neck squamous cell carcinoma; AML: acute myelocytic leukemia.
Acknowledgements
Funding
This study was funded by the State Key Project on Infection Disease of China (No. 2018ZX10723204-003); the Chinese Ministry of Public Health for Key Clinical Project (No. [2010] 493-51); National Natural Science Foundation of China (81874189).
Author contributions
MZ Han collected the related papers and was a major contributor in writing the manuscript. QB Huang and XX Li generated the data and prepared the figures. XP Chen provided the advices of this study. H Zhu, YL Pan and BX Zhang initiated the study and revised the manuscript. BX Zhang provided financial and administrative support. All authors read and approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Zhang Y, Lu L, Li X. Detection technologies for RNA modifications. Exp Mol Med. 2022;54:1601-16
2. Huang H, Weng H, Chen J. m(6)A Modification in Coding and Non-coding RNAs: Roles and Therapeutic Implications in Cancer. Cancer Cell. 2020;37:270-88
3. Han D, Xu MM. RNA Modification in the Immune System. Annu Rev Immunol. 2023;41:73-98
4. Xue C, Chu Q, Zheng Q, Jiang S, Bao Z, Su Y. et al. Role of main RNA modifications in cancer: N(6)-methyladenosine, 5-methylcytosine, and pseudouridine. Signal Transduct Tar. 2022;7:142
5. Yang J, Xu J, Wang W, Zhang B, Yu X, Shi S. Epigenetic regulation in the tumor microenvironment: molecular mechanisms and therapeutic targets. Signal Transduct Tar. 2023;8:210
6. Cui L, Ma R, Cai J, Guo C, Chen Z, Yao L. et al. RNA modifications: importance in immune cell biology and related diseases. Signal Transduct Tar. 2022;7:334
7. Yang Y, Hsu PJ, Chen YS, Yang YG. Dynamic transcriptomic m(6)A decoration: writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018;28:616-24
8. He C, Bozler J, Janssen KA, Wilusz JE, Garcia BA, Schorn AJ. et al. TET2 chemically modifies tRNAs and regulates tRNA fragment levels. Nat Struct Mol Biol. 2021;28:62-70
9. Karijolich J, Yi C, Yu YT. Transcriptome-wide dynamics of RNA pseudouridylation. Nat Rev Mol Cell Bio. 2015;16:581-5
10. Chen Y, Lin Y, Shu Y, He J, Gao W. Interaction between N(6)-methyladenosine (m(6)A) modification and noncoding RNAs in cancer. Mol Cancer. 2020;19:94
11. Yi YC, Chen XY, Zhang J, Zhu JS. Novel insights into the interplay between m(6)A modification and noncoding RNAs in cancer. Mol Cancer. 2020;19:121
12. Sun H, Li K, Liu C, Yi C. Regulation and functions of non-m(6)A mRNA modifications. Nat Rev Mol Cell Bio. 2023
13. Zhang M, Song J, Yuan W, Zhang W, Sun Z. Roles of RNA Methylation on Tumor Immunity and Clinical Implications. Front Immunol. 2021;12:641507
14. Feng Q, Wang D, Xue T, Lin C, Gao Y, Sun L. et al. The role of RNA modification in hepatocellular carcinoma. Front Pharmacol. 2022;13:984453
15. Xia X, Wang Y, Zheng JC. Internal m7G methylation: A novel epitranscriptomic contributor in brain development and diseases. Mol Ther-Nucl Acids. 2023;31:295-308
16. Cai M, Yang C, Wang Z. N7-methylguanosine modification: from regulatory roles to therapeutic implications in cancer. Am J Cancer Res. 2023;13:1640-55
17. Malbec L, Zhang T, Chen YS, Zhang Y, Sun BF, Shi BY. et al. Dynamic methylome of internal mRNA N(7)-methylguanosine and its regulatory role in translation. Cell Res. 2019;29:927-41
18. Alexandrov A, Martzen MR, Phizicky EM. Two proteins that form a complex are required for 7-methylguanosine modification of yeast tRNA. Rna. 2002;8:1253-66
19. Zhang LS, Liu C, Ma H, Dai Q, Sun HL, Luo G. et al. Transcriptome-wide Mapping of Internal N(7)-Methylguanosine Methylome in Mammalian mRNA. Mol Cell. 2019;74:1304-16
20. Lin S, Liu Q, Jiang YZ, Gregory RI. Nucleotide resolution profiling of m(7)G tRNA modification by TRAC-Seq. Nat Protoc. 2019;14:3220-42
21. Gatsiou A, Stellos K. RNA modifications in cardiovascular health and disease. Nat Rev Cardiol. 2023;20:325-46
22. Hanahan D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022;12:31-46
23. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646-74
24. Duan Q, Zhang H, Zheng J, Zhang L. Turning Cold into Hot: Firing up the Tumor Microenvironment. Trends Cancer. 2020;6:605-18
25. Barbieri I, Kouzarides T. Role of RNA modifications in cancer. Nat Rev Cancer. 2020;20:303-22
26. Han X, Wang M, Zhao YL, Yang Y, Yang YG. RNA methylations in human cancers. Semin Cancer Biol. 2021;75:97-115
27. Alexandrov A, Grayhack EJ, Phizicky EM. tRNA m7G methyltransferase Trm8p/Trm82p: evidence linking activity to a growth phenotype and implicating Trm82p in maintaining levels of active Trm8p. Rna. 2005;11:821-30
28. Tang Q, Li L, Wang Y, Wu P, Hou X, Ouyang J. et al. RNA modifications in cancer. Brit J Cancer. 2023;129:204-21
29. Pisera A, Campo A, Campo S. Structure and functions of the translation initiation factor eIF4E and its role in cancer development and treatment. J Genet Genomics. 2018;45:13-24
30. Cowling VH. Regulation of mRNA cap methylation. Biochem J. 2009;425:295-302
31. Gonatopoulos-Pournatzis T, Dunn S, Bounds R, Cowling VH. RAM/Fam103a1 is required for mRNA cap methylation. Mol Cell. 2011;44:585-96
32. Varshney D, Petit AP, Bueren-Calabuig JA, Jansen C, Fletcher DA, Peggie M. et al. Molecular basis of RNA guanine-7 methyltransferase (RNMT) activation by RAM. Nucleic Acids Res. 2016;44:10423-36
33. Bueren-Calabuig JA, G BM, Cowling VH, Pisliakov AV. Mechanism of allosteric activation of human mRNA cap methyltransferase (RNMT) by RAM: insights from accelerated molecular dynamics simulations. Nucleic Acids Res. 2019;47:8675-92
34. Rambout X, Maquat LE. The nuclear cap-binding complex as choreographer of gene transcription and pre-mRNA processing. Gene Dev. 2020;34:1113-27
35. Cowling VH. Enhanced mRNA cap methylation increases cyclin D1 expression and promotes cell transformation. Oncogene. 2010;29:930-6
36. Posternak V, Ung MH, Cheng C, Cole MD. MYC Mediates mRNA Cap Methylation of Canonical Wnt/beta-Catenin Signaling Transcripts By Recruiting CDK7 and RNA Methyltransferase. Mol Cancer Res. 2017;15:213-24
37. Aregger M, Kaskar A, Varshney D, Fernandez-Sanchez ME, Inesta-Vaquera FA, Weidlich S. et al. CDK1-Cyclin B1 Activates RNMT, Coordinating mRNA Cap Methylation with G1 Phase Transcription. Mol Cell. 2016;61:734-46
38. Osborne MJ, Volpon L, Memarpoor-Yazdi M, Pillay S, Thambipillai A, Czarnota S. et al. Identification and Characterization of the Interaction Between the Methyl-7-Guanosine Cap Maturation Enzyme RNMT and the Cap-Binding Protein eIF4E. J Mol Biol. 2022;434:167451
39. Lin S, Liu Q, Lelyveld VS, Choe J, Szostak JW, Gregory RI. Mettl1/Wdr4-Mediated m(7)G tRNA Methylome Is Required for Normal mRNA Translation and Embryonic Stem Cell Self-Renewal and Differentiation. Mol Cell. 2018;71:244-55
40. Ruiz-Arroyo VM, Raj R, Babu K, Onolbaatar O, Roberts PH, Nam Y. Structures and mechanisms of tRNA methylation by METTL1-WDR4. Nature. 2023;613:383-90
41. Li J, Wang L, Hahn Q, Nowak RP, Viennet T, Orellana EA. et al. Structural basis of regulated m(7)G tRNA modification by METTL1-WDR4. Nature. 2023;613:391-7
42. Cartlidge RA, Knebel A, Peggie M, Alexandrov A, Phizicky EM, Cohen P. The tRNA methylase METTL1 is phosphorylated and inactivated by PKB and RSK in vitro and in cells. Embo J. 2005;24:1696-705
43. Dedon PC, Begley TJ. Dysfunctional tRNA reprogramming and codon-biased translation in cancer. Trends Mol Med. 2022;28:964-78
44. Katsara O, Schneider RJ. m(7)G tRNA modification reveals new secrets in the translational regulation of cancer development. Mol Cell. 2021;81:3243-5
45. Tomikawa C. 7-Methylguanosine Modifications in Transfer RNA (tRNA). Int J Mol Sci. 2018 19
46. Ma J, Han H, Huang Y, Yang C, Zheng S, Cai T. et al. METTL1/WDR4-mediated m(7)G tRNA modifications and m(7)G codon usage promote mRNA translation and lung cancer progression. Mol Ther. 2021;29:3422-35
47. Dai Z, Liu H, Liao J, Huang C, Ren X, Zhu W. et al. N(7)-Methylguanosine tRNA modification enhances oncogenic mRNA translation and promotes intrahepatic cholangiocarcinoma progression. Mol Cell. 2021;81:3339-55
48. Chen Z, Zhu W, Zhu S, Sun K, Liao J, Liu H. et al. METTL1 promotes hepatocarcinogenesis via m(7) G tRNA modification-dependent translation control. Clin Transl Med. 2021;11:e661
49. Han H, Yang C, Ma J, Zhang S, Zheng S, Ling R. et al. N(7)-methylguanosine tRNA modification promotes esophageal squamous cell carcinoma tumorigenesis via the RPTOR/ULK1/autophagy axis. Nat Commun. 2022;13:1478
50. Orellana EA, Liu Q, Yankova E, Pirouz M, De Braekeleer E, Zhang W. et al. METTL1-mediated m(7)G modification of Arg-TCT tRNA drives oncogenic transformation. Mol Cell. 2021;81:3323-38
51. Cheng W, Gao A, Lin H, Zhang W. Novel roles of METTL1/WDR4 in tumor via m(7)G methylation. Mol Ther-Oncolytics. 2022;26:27-34
52. Zhao Z, Qing Y, Dong L, Han L, Wu D, Li Y. et al. QKI shuttles internal m(7)G-modified transcripts into stress granules and modulates mRNA metabolism. Cell. 2023;186:3208-26
53. Zhang M, Kan D, Zhang B, Chen X, Wang C, Chen S. et al. P300/SP1 complex mediating elevated METTL1 regulates CDK14 mRNA stability via internal m7G modification in CRPC. J Exp Clin Canc Res. 2023;42:215
54. Pandolfini L, Barbieri I, Bannister AJ, Hendrick A, Andrews B, Webster N. et al. METTL1 Promotes let-7 MicroRNA Processing via m7G Methylation. Mol Cell. 2019;74:1278-90
55. Xie H, Wang M, Yu H, Wang H, Ding L, Wang R. et al. METTL1 drives tumor progression of bladder cancer via degrading ATF3 mRNA in an m(7)G-modified miR-760-dependent manner. Cell Death Discov. 2022;8:458
56. Rong J, Wang H, Yao Y, Wu Z, Chen L, Jin C. et al. Identification of m7G-associated lncRNA prognostic signature for predicting the immune status in cutaneous melanoma. Aging (Albany NY). 2022;14:5233-49
57. Nakazawa Y, Arai H, Fujita N. The novel metastasis promoter Merm1/Wbscr22 enhances tumor cell survival in the vasculature by suppressing Zac1/p53-dependent apoptosis. Cancer Res. 2011;71:1146-55
58. Haag S, Kretschmer J, Bohnsack MT. WBSCR22/Merm1 is required for late nuclear pre-ribosomal RNA processing and mediates N7-methylation of G1639 in human 18S rRNA. Rna. 2015;21:180-7
59. Letoquart J, Huvelle E, Wacheul L, Bourgeois G, Zorbas C, Graille M. et al. Structural and functional studies of Bud23-Trm112 reveal 18S rRNA N7-G1575 methylation occurs on late 40S precursor ribosomes. P Natl Acad Sci Usa. 2014;111:E5518-26
60. Ounap K, Kasper L, Kurg A, Kurg R. The human WBSCR22 protein is involved in the biogenesis of the 40S ribosomal subunits in mammalian cells. Plos One. 2013;8:e75686
61. Brumele B, Mutso M, Telanne L, Ounap K, Spunde K, Abroi A. et al. Human TRMT112-Methyltransferase Network Consists of Seven Partners Interacting with a Common Co-Factor. Int J Mol Sci. 2021 22
62. Yan D, Tu L, Yuan H, Fang J, Cheng L, Zheng X. et al. WBSCR22 confers oxaliplatin resistance in human colorectal cancer. Sci Rep-Uk. 2017;7:15443
63. Khan AA, Huang H, Zhao Y, Li H, Pan R, Wang S. et al. WBSCR22 and TRMT112 synergistically suppress cell proliferation, invasion and tumorigenesis in pancreatic cancer via transcriptional regulation of ISG15. Int J Oncol. 2022 60
64. Mars JC, Ghram M, Culjkovic-Kraljacic B, Borden K. The Cap-Binding Complex CBC and the Eukaryotic Translation Factor eIF4E: Co-Conspirators in Cap-Dependent RNA Maturation and Translation. Cancers. 2021 13
65. Gonatopoulos-Pournatzis T, Cowling VH. Cap-binding complex (CBC). Biochem J. 2014;457:231-42
66. Borden K, Culjkovic-Kraljacic B, Cowling VH. To cap it all off, again: dynamic capping and recapping of coding and non-coding RNAs to control transcript fate and biological activity. Cell Cycle. 2021;20:1347-60
67. Xie M, Li M, Vilborg A, Lee N, Shu MD, Yartseva V. et al. Mammalian 5'-capped microRNA precursors that generate a single microRNA. Cell. 2013;155:1568-80
68. Culjkovic-Kraljacic B, Skrabanek L, Revuelta MV, Gasiorek J, Cowling VH, Cerchietti L. et al. The eukaryotic translation initiation factor eIF4E elevates steady-state m(7)G capping of coding and noncoding transcripts. P Natl Acad Sci Usa. 2020;117:26773-83
69. Volpon L, Culjkovic-Kraljacic B, Osborne MJ, Ramteke A, Sun Q, Niesman A. et al. Importin 8 mediates m7G cap-sensitive nuclear import of the eukaryotic translation initiation factor eIF4E. P Natl Acad Sci Usa. 2016;113:5263-8
70. Zucko D, Boris-Lawrie K. Blocking tri-methylguanosine synthase 1 (TGS1) stops anchorage-independent growth of canine sarcomas. Cancer Gene Ther. 2023
71. Culjkovic B, Topisirovic I, Skrabanek L, Ruiz-Gutierrez M, Borden KL. eIF4E promotes nuclear export of cyclin D1 mRNAs via an element in the 3'UTR. J Cell Biol. 2005;169:245-56
72. Osborne MJ, Volpon L, Kornblatt JA, Culjkovic-Kraljacic B, Baguet A, Borden KL. eIF4E3 acts as a tumor suppressor by utilizing an atypical mode of methyl-7-guanosine cap recognition. P Natl Acad Sci Usa. 2013;110:3877-82
73. Chen B, Jiang W, Huang Y, Zhang J, Yu P, Wu L. et al. N(7)-methylguanosine tRNA modification promotes tumorigenesis and chemoresistance through WNT/beta-catenin pathway in nasopharyngeal carcinoma. Oncogene. 2022;41:2239-53
74. Xia P, Zhang H, Xu K, Jiang X, Gao M, Wang G. et al. MYC-targeted WDR4 promotes proliferation, metastasis, and sorafenib resistance by inducing CCNB1 translation in hepatocellular carcinoma. Cell Death Dis. 2021;12:691
75. Huang X, Chen Z, Xiang X, Liu Y, Long X, Li K. et al. Comprehensive multi-omics analysis of the m7G in pan-cancer from the perspective of predictive, preventive, and personalized medicine. Epma J. 2022;13:671-97
76. Zeng X, Liao G, Li S, Liu H, Zhao X, Li S. et al. Eliminating METTL1-mediated accumulation of PMN-MDSCs prevents hepatocellular carcinoma recurrence after radiofrequency ablation. Hepatology. 2023;77:1122-38
77. Yang L, Chu Z, Liu M, Zou Q, Li J, Liu Q. et al. Amino acid metabolism in immune cells: essential regulators of the effector functions, and promising opportunities to enhance cancer immunotherapy. J Hematol Oncol. 2023;16:59
78. Hiam-Galvez KJ, Allen BM, Spitzer MH. Systemic immunity in cancer. Nat Rev Cancer. 2021;21:345-59
79. Kim M, Lee NK, Wang CJ, Lim J, Byun MJ, Kim TH. et al. Reprogramming the tumor microenvironment with biotechnology. Biomater Res. 2023;27:5
80. Sun L, Su Y, Jiao A, Wang X, Zhang B. T cells in health and disease. Signal Transduct Tar. 2023;8:235
81. Bertolini M, Wong MS, Mendive-Tapia L, Vendrell M. Smart probes for optical imaging of T cells and screening of anti-cancer immunotherapies. Chem Soc Rev. 2023;52:5352-72
82. Peri A, Salomon N, Wolf Y, Kreiter S, Diken M, Samuels Y. The landscape of T cell antigens for cancer immunotherapy. Nat Cancer. 2023;4:937-54
83. Wang R, Liu Z, Fan Z, Zhan H. Lipid metabolism reprogramming of CD8(+) T cell and therapeutic implications in cancer. Cancer Lett. 2023;567:216267
84. Cao J, Liao S, Zeng F, Liao Q, Luo G, Zhou Y. Effects of altered glycolysis levels on CD8(+) T cell activation and function. Cell Death Dis. 2023;14:407
85. Xu X, Zhao Y, Ying Y, Zhu H, Luo J, Mou T. et al. m7G-related genes-NCBP2 and EIF4E3 determine immune contexture in head and neck squamous cell carcinoma by regulating CCL4/CCL5 expression. Mol Carcinogen. 2023;62:1091-106
86. Qi L, Zhang W, Ren X, Xu R, Yang Z, Chen R. et al. Cross-Talk of Multiple Types of RNA Modification Regulators Uncovers the Tumor Microenvironment and Immune Infiltrates in Soft Tissue Sarcoma. Front Immunol. 2022;13:921223
87. Cheng Z, Wang J, Xu Y, Jiang T, Xue Z, Li S. et al. N7-methylguanosine-related lncRNAs: Distinction between hot and cold tumors and construction of predictive models in colon adenocarcinoma. Front Oncol. 2022;12:951452
88. Li XY, Wang SL, Chen DH, Liu H, You JX, Su LX. et al. Construction and Validation of a m7G-Related Gene-Based Prognostic Model for Gastric Cancer. Front Oncol. 2022;12:861412
89. Xin S, Deng Y, Mao J, Wang T, Liu J, Wang S. et al. Characterization of 7-Methylguanosine Identified Biochemical Recurrence and Tumor Immune Microenvironment in Prostate Cancer. Front Oncol. 2022;12:900203
90. Gao K, Lian W, Zhao R, Huang W, Xiong J. The joint role of methylation and immune-related lncRNAs in ovarian cancer: Defining molecular subtypes and developing prognostic signature. Transl Oncol. 2023;34:101704
91. Taniuchi I. CD4 Helper and CD8 Cytotoxic T Cell Differentiation. Annu Rev Immunol. 2018;36:579-601
92. Geginat J, Paroni M, Maglie S, Alfen JS, Kastirr I, Gruarin P. et al. Plasticity of human CD4 T cell subsets. Front Immunol. 2014;5:630
93. Walker JA, McKenzie A. T(H)2 cell development and function. Nat Rev Immunol. 2018;18:121-33
94. Maharaj K, Uriepero A, Sahakian E, Pinilla-Ibarz J. Regulatory T cells (Tregs) in lymphoid malignancies and the impact of novel therapies. Front Immunol. 2022;13:943354
95. Vinay DS, Ryan EP, Pawelec G, Talib WH, Stagg J, Elkord E. et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol. 2015;35(Suppl):S185-98
96. Huang X, Zhu B, Qian C, Feng Y. The prognostic index of m(7)G-related genes in CRC correlates with immune infiltration. Sci Rep-Uk. 2022;12:21282
97. Gao Z, Xu J, Zhang Z, Fan Y, Xue H, Guo X. et al. A Comprehensive Analysis of METTL1 to Immunity and Stemness in Pan-Cancer. Front Immunol. 2022;13:795240
98. Wang YT, Chen J, Chang CW, Jen J, Huang TY, Chen CM. et al. Ubiquitination of tumor suppressor PML regulates prometastatic and immunosuppressive tumor microenvironment. J Clin Invest. 2017;127:2982-97
99. Chen J, Li K, Chen J, Wang X, Ling R, Cheng M. et al. Aberrant translation regulated by METTL1/WDR4-mediated tRNA N7-methylguanosine modification drives head and neck squamous cell carcinoma progression. Cancer Commun. 2022;42:223-44
100. Galloway A, Kaskar A, Ditsova D, Atrih A, Yoshikawa H, Gomez-Moreira C. et al. Upregulation of RNA cap methyltransferase RNMT drives ribosome biogenesis during T cell activation. Nucleic Acids Res. 2021;49:6722-38
101. Jangani M, Poolman TM, Matthews L, Yang N, Farrow SN, Berry A. et al. The methyltransferase WBSCR22/Merm1 enhances glucocorticoid receptor function and is regulated in lung inflammation and cancer. J Biol Chem. 2014;289:8931-46
102. Hu X, Zhang J, Wang J, Fu J, Li T, Zheng X. et al. Landscape of B cell immunity and related immune evasion in human cancers. Nat Genet. 2019;51:560-7
103. Laumont CM, Nelson BH. B cells in the tumor microenvironment: Multi-faceted organizers, regulators, and effectors of anti-tumor immunity. Cancer Cell. 2023;41:466-89
104. Jiang S, Xiao M, Shi Y, Wang Y, Xu Z, Wang K. Identification of m7G-Related miRNA Signatures Associated with Prognosis, Oxidative Stress, and Immune Landscape in Lung Adenocarcinoma. Biomedicines. 2023 11
105. Chen J, Song YW, Liang GZ, Zhang ZJ, Wen XF, Li RB. et al. A Novel m7G-Related Gene Signature Predicts the Prognosis of Colon Cancer. Cancers. 2022 14
106. Zhang M, Yang L, Wang Y, Zuo Y, Chen D, Guo X. Comprehensive prediction of immune microenvironment and hot and cold tumor differentiation in cutaneous melanoma based on necroptosis-related lncRNA. Sci Rep-Uk. 2023;13:7299
107. Li DX, Feng DC, Wang XM, Wu RC, Zhu WZ, Chen K. et al. M7G-related molecular subtypes can predict the prognosis and correlate with immunotherapy and chemotherapy responses in bladder cancer patients. Eur J Med Res. 2023;28:55
108. Wang T, Zhou Z, Wang X, You L, Li W, Zheng C. et al. Comprehensive analysis of nine m7G-related lncRNAs as prognosis factors in tumor immune microenvironment of hepatocellular carcinoma and experimental validation. Front Genet. 2022;13:929035
109. Li X, Dong H, Chen L, Wang Y, Hao Z, Zhang Y. et al. Identification of N7-methylguanosine related subtypes and construction of prognostic model in gastric cancer. Front Immunol. 2022;13:984149
110. Hegde S, Leader AM, Merad M. MDSC: Markers, development, states, and unaddressed complexity. Immunity. 2021;54:875-84
111. Wu Y, Yi M, Niu M, Mei Q, Wu K. Myeloid-derived suppressor cells: an emerging target for anticancer immunotherapy. Mol Cancer. 2022;21:184
112. Liu H, Zeng X, Ren X, Zhang Y, Huang M, Tan L. et al. Targeting tumour-intrinsic N(7)-methylguanosine tRNA modification inhibits MDSC recruitment and improves anti-PD-1 efficacy. Gut. 2023;72:1555-67
113. Yang F, Jiang S, Zheng Y. Letter to the Editor: Is eliminating METTL1-mediated accumulation of PMN-MDSCs sufficient to prevent hepatocellular carcinoma recurrence after radiofrequency ablation? Hepatology. 2023;77:E62-3
114. Zhang X, Li Y, Hu P, Xu L, Qiu H. KIF2C is a Biomarker Correlated With Prognosis and Immunosuppressive Microenvironment in Human Tumors. Front Genet. 2022;13:891408
115. Wang L, Hu X, Liu X, Feng Y, Zhang Y, Han J. et al. m(7)G regulator-mediated methylation modification patterns define immune cell infiltration and patient survival. Front Immunol. 2022;13:1022720
116. Zhang X, Miao Y, Sun HW, Wang YX, Zhao WM, Pang AY. et al. Integrated analysis from multi-center studies identities m7G-derived modification pattern and risk stratification system in skin cutaneous melanoma. Front Immunol. 2022;13:1034516
117. Li M, Yang Y, Xiong L, Jiang P, Wang J, Li C. Metabolism, metabolites, and macrophages in cancer. J Hematol Oncol. 2023;16:80
118. Bied M, Ho WW, Ginhoux F, Bleriot C. Roles of macrophages in tumor development: a spatiotemporal perspective. Cell Mol Immunol. 2023
119. Chen S, Saeed A, Liu Q, Jiang Q, Xu H, Xiao GG. et al. Macrophages in immunoregulation and therapeutics. Signal Transduct Tar. 2023;8:207
120. Iglesias-Escudero M, Arias-Gonzalez N, Martinez-Caceres E. Regulatory cells and the effect of cancer immunotherapy. Mol Cancer. 2023;22:26
121. Wang J, Mi S, Ding M, Li X, Yuan S. Metabolism and polarization regulation of macrophages in the tumor microenvironment. Cancer Lett. 2022;543:215766
122. Xu Z, Chen Y, Ma L, Chen Y, Liu J, Guo Y. et al. Role of exosomal non-coding RNAs from tumor cells and tumor-associated macrophages in the tumor microenvironment. Mol Ther. 2022;30:3133-54
123. Garcia-Vilchez R, Anazco-Guenkova AM, Dietmann S, Lopez J, Moron-Calvente V, D'Ambrosi S. et al. METTL1 promotes tumorigenesis through tRNA-derived fragment biogenesis in prostate cancer. Mol Cancer. 2023;22:119
124. Xu F, Cai D, Liu S, He K, Chen J, Qu L. et al. N7-methylguanosine regulatory genes well represented by METTL1 define vastly different prognostic, immune and therapy landscapes in adrenocortical carcinoma. Am J Cancer Res. 2023;13:538-68
125. Yang H, Messina-Pacheco J, Corredor A, Gregorieff A, Liu JL, Nehme A. et al. An integrated model of acinar to ductal metaplasia-related N7-methyladenosine regulators predicts prognosis and immunotherapy in pancreatic carcinoma based on digital spatial profiling. Front Immunol. 2022;13:961457
126. Zheng P, Li N, Zhan X. Ovarian cancer subtypes based on the regulatory genes of RNA modifications: Novel prediction model of prognosis. Front Endocrinol. 2022;13:972341
127. Ren L, Yang X, Liu J, Wang W, Liu Z, Lin Q. et al. An innovative model based on N7-methylguanosine-related lncRNAs for forecasting prognosis and tumor immune landscape in bladder cancer. Cancer Cell Int. 2023;23:85
128. Yang S, Zhou J, Chen Z, Sun Q, Zhang D, Feng Y. et al. A novel m7G-related lncRNA risk model for predicting prognosis and evaluating the tumor immune microenvironment in colon carcinoma. Front Oncol. 2022;12:934928
129. Liu S, Wu W, Du Y, Yin H, Chen Q, Yu W. et al. The evolution and heterogeneity of neutrophils in cancers: origins, subsets, functions, orchestrations and clinical applications. Mol Cancer. 2023;22:148
130. Adrover JM, McDowell S, He XY, Quail DF, Egeblad M. NETworking with cancer: The bidirectional interplay between cancer and neutrophil extracellular traps. Cancer Cell. 2023;41:505-26
131. Dutta A, Bhagat S, Paul S, Katz JP, Sengupta D, Bhargava D. Neutrophils in Cancer and Potential Therapeutic Strategies Using Neutrophil-Derived Exosomes. Vaccines-Basel. 2023 11
132. Zhang W, Zhang S, Wang Z. Prognostic value of 12 m7G methylation-related miRNA markers and their correlation with immune infiltration in breast cancer. Front Oncol. 2022;12:929363
133. Dong Y, Li Y, Yao Y, Song Q. A novel defined m7G regulator signature to investigate the association between molecular characterization and clinical significance in lung adenocarcinoma. Front Oncol. 2022;12:897323
134. Ran GH, Lin YQ, Tian L, Zhang T, Yan DM, Yu JH. et al. Natural killer cell homing and trafficking in tissues and tumors: from biology to application. Signal Transduct Tar. 2022;7:205
135. Del PA, Salvi V, Soriani A, Laffranchi M, Sozio F, Bosisio D. et al. Dendritic cell subsets in cancer immunity and tumor antigen sensing. Cell Mol Immunol. 2023;20:432-47
136. Pittet MJ, Di Pilato M, Garris C, Mempel TR. Dendritic cells as shepherds of T cell immunity in cancer. Immunity. 2023
137. Komi D, Redegeld FA. Role of Mast Cells in Shaping the Tumor Microenvironment. Clin Rev Allerg Immu. 2020;58:313-25
138. Li Z, Wang W, Wu J, Ye X. Identification of N7-methylguanosine related signature for prognosis and immunotherapy efficacy prediction in lung adenocarcinoma. Front Med-Lausanne. 2022;9:962972
139. Mao X, Xu J, Wang W, Liang C, Hua J, Liu J. et al. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Mol Cancer. 2021;20:131
140. Wilky BA. Immune checkpoint inhibitors: The linchpins of modern immunotherapy. Immunol Rev. 2019;290:6-23
141. Qian W, Zhao M, Wang R, Li H. Fibrinogen-like protein 1 (FGL1): the next immune checkpoint target. J Hematol Oncol. 2021;14:147
142. Pathania AS, Prathipati P, Murakonda SP, Murakonda AB, Srivastava A, Avadhesh. et al. Immune checkpoint molecules in neuroblastoma: A clinical perspective. Semin Cancer Biol. 2022;86:247-58
143. Wang Y, Zhang H, Liu C, Wang Z, Wu W, Zhang N. et al. Immune checkpoint modulators in cancer immunotherapy: recent advances and emerging concepts. J Hematol Oncol. 2022;15:111
144. Sun Q, Hong Z, Zhang C, Wang L, Han Z, Ma D. Immune checkpoint therapy for solid tumours: clinical dilemmas and future trends. Signal Transduct Tar. 2023;8:320
145. Ma M, Li J, Zeng Z, Zheng Z, Kang W. Integrated analysis from multicentre studies identities m7G-related lncRNA-derived molecular subtypes and risk stratification systems for gastric cancer. Front Immunol. 2023;14:1096488
146. Maimaiti A, Feng Z, Liu Y, Turhon M, Xie Z, Baihetiyaer Y. et al. N7-methylguanosin regulators-mediated methylation modification patterns and characterization of the immune microenvironment in lower-grade glioma. Eur J Med Res. 2023;28:144
147. Li K, Wang W. Establishment of m7G-related gene pair signature to predict overall survival in colorectal cancer. Front Genet. 2022;13:981392
148. de Visser KE, Joyce JA. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell. 2023;41:374-403
149. Qu P, Wang LZ, Lin PC. Expansion and functions of myeloid-derived suppressor cells in the tumor microenvironment. Cancer Lett. 2016;380:253-6
150. Turley SJ, Cremasco V, Astarita JL. Immunological hallmarks of stromal cells in the tumour microenvironment. Nat Rev Immunol. 2015;15:669-82
151. Zhao F, Dong Z, Li Y, Liu S, Guo P, Zhang D. et al. Comprehensive Analysis of Molecular Clusters and Prognostic Signature Based on m7G-related LncRNAs in Esophageal Squamous Cell Carcinoma. Front Oncol. 2022;12:893186
152. Lu F, Gao J, Hou Y, Cao K, Xia Y, Chen Z. et al. Construction of a Novel Prognostic Model in Lung Adenocarcinoma Based on 7-Methylguanosine-Related Gene Signatures. Front Oncol. 2022;12:876360
153. Zhao Y, Kong L, Pei Z, Li F, Li C, Sun X. et al. m7G Methyltransferase METTL1 Promotes Post-ischemic Angiogenesis via Promoting VEGFA mRNA Translation. Front Cell Dev Biol. 2021;9:642080
154. Qu X, Zhou D, Lu J, Qin D, Zhou J, Liu HJ. Cancer nanomedicine in preoperative therapeutics: Nanotechnology-enabled neoadjuvant chemotherapy, radiotherapy, immunotherapy, and phototherapy. Bioact Mater. 2023;24:136-52
155. Sharma P, Goswami S, Raychaudhuri D, Siddiqui BA, Singh P, Nagarajan A. et al. Immune checkpoint therapy-current perspectives and future directions. Cell. 2023;186:1652-69
156. Wolchok JD, Neyns B, Linette G, Negrier S, Lutzky J, Thomas L. et al. Ipilimumab monotherapy in patients with pretreated advanced melanoma: a randomised, double-blind, multicentre, phase 2, dose-ranging study. Lancet Oncol. 2010;11:155-64
157. Livingstone E, Zimmer L, Hassel JC, Fluck M, Eigentler TK, Loquai C. et al. Adjuvant nivolumab plus ipilimumab or nivolumab alone versus placebo in patients with resected stage IV melanoma with no evidence of disease (IMMUNED): final results of a randomised, double-blind, phase 2 trial. Lancet. 2022;400:1117-29
158. Schachter J, Ribas A, Long GV, Arance A, Grob JJ, Mortier L. et al. Pembrolizumab versus ipilimumab for advanced melanoma: final overall survival results of a multicentre, randomised, open-label phase 3 study (KEYNOTE-006). Lancet. 2017;390:1853-62
159. Kwon ED, Drake CG, Scher HI, Fizazi K, Bossi A, van den Eertwegh AJ. et al. Ipilimumab versus placebo after radiotherapy in patients with metastatic castration-resistant prostate cancer that had progressed after docetaxel chemotherapy (CA184-043): a multicentre, randomised, double-blind, phase 3 trial. Lancet Oncol. 2014;15:700-12
160. Scott EC, Baines AC, Gong Y, Moore RJ, Pamuk GE, Saber H. et al. Trends in the approval of cancer therapies by the FDA in the twenty-first century. Nat Rev Drug Discov. 2023;22:625-40
161. Zimmer L, Livingstone E, Hassel JC, Fluck M, Eigentler T, Loquai C. et al. Adjuvant nivolumab plus ipilimumab or nivolumab monotherapy versus placebo in patients with resected stage IV melanoma with no evidence of disease (IMMUNED): a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet. 2020;395:1558-68
162. Cortes J, Cescon DW, Rugo HS, Nowecki Z, Im SA, Yusof MM. et al. Pembrolizumab plus chemotherapy versus placebo plus chemotherapy for previously untreated locally recurrent inoperable or metastatic triple-negative breast cancer (KEYNOTE-355): a randomised, placebo-controlled, double-blind, phase 3 clinical trial. Lancet. 2020;396:1817-28
163. Hellmann MD, Paz-Ares L, Bernabe CR, Zurawski B, Kim SW, Carcereny CE. et al. Nivolumab plus Ipilimumab in Advanced Non-Small-Cell Lung Cancer. New Engl J Med. 2019;381:2020-31
164. Gao X, Xu N, Li Z, Shen L, Ji K, Zheng Z. et al. Safety and antitumour activity of cadonilimab, an anti-PD-1/CTLA-4 bispecific antibody, for patients with advanced solid tumours (COMPASSION-03): a multicentre, open-label, phase 1b/2 trial. Lancet Oncol. 2023;24:1134-46
165. Luke JJ, Patel MR, Blumenschein GR, Hamilton E, Chmielowski B, Ulahannan SV. et al. The PD-1- and LAG-3-targeting bispecific molecule tebotelimab in solid tumors and hematologic cancers: a phase 1 trial. Nat Med. 2023;29:2814-24
166. Xiong A, Li W, Li X, Fan Y, Ma Z, Fang J. et al. Efficacy and safety of KN046, a novel bispecific antibody against PD-L1 and CTLA-4, in patients with non-small cell lung cancer who failed platinum-based chemotherapy: a phase II study. Eur J Cancer. 2023;190:112936
167. Jardim DL, Goodman A, de Melo GD, Kurzrock R. The Challenges of Tumor Mutational Burden as an Immunotherapy Biomarker. Cancer Cell. 2021;39:154-73
168. Jiang P, Gu S, Pan D, Fu J, Sahu A, Hu X. et al. Signatures of T cell dysfunction and exclusion predict cancer immunotherapy response. Nat Med. 2018;24:1550-8
169. Li Z, Zhao J, Huang X, Wang J. An m7G-related lncRNA signature predicts prognosis and reveals the immune microenvironment in bladder cancer. Sci Rep-Uk. 2023;13:4302
170. Lai G, Zhong X, Liu H, Deng J, Li K, Xie B. A Novel m7G-Related Genes-Based Signature with Prognostic Value and Predictive Ability to Select Patients Responsive to Personalized Treatment Strategies in Bladder Cancer. Cancers. 2022 14
171. Wang Z, Yu P, Zou Y, Ma J, Han H, Wei W. et al. METTL1/WDR4-mediated tRNA m(7)G modification and mRNA translation control promote oncogenesis and doxorubicin resistance. Oncogene. 2023;42:1900-12
172. Assouline S, Culjkovic B, Cocolakis E, Rousseau C, Beslu N, Amri A. et al. Molecular targeting of the oncogene eIF4E in acute myeloid leukemia (AML): a proof-of-principle clinical trial with ribavirin. Blood. 2009;114:257-60
173. Boussemart L, Malka-Mahieu H, Girault I, Allard D, Hemmingsson O, Tomasic G. et al. eIF4F is a nexus of resistance to anti-BRAF and anti-MEK cancer therapies. Nature. 2014;513:105-9
174. Han WY, Wang J, Zhao J, Zheng YM, Chai XQ, Gao C. et al. WDR4/TRIM28 is a novel molecular target linked to lenvatinib resistance that helps retain the stem characteristics in hepatocellular carcinomas. Cancer Lett. 2023;568:216259
175. Huang M, Long J, Yao Z, Zhao Y, Zhao Y, Liao J. et al. METTL1-Mediated m7G tRNA Modification Promotes Lenvatinib Resistance in Hepatocellular Carcinoma. Cancer Res. 2023;83:89-102
176. Liao J, Yi Y, Yue X, Wu X, Zhu M, Chen Y. et al. Methyltransferase 1 is required for nonhomologous end-joining repair and renders hepatocellular carcinoma resistant to radiotherapy. Hepatology. 2023;77:1896-910
177. Zhu S, Wu Y, Zhang X, Peng S, Xiao H, Chen S. et al. Targeting N(7)-methylguanosine tRNA modification blocks hepatocellular carcinoma metastasis after insufficient radiofrequency ablation. Mol Ther. 2023;31:1596-614
178. Dunn S, Lombardi O, Lukoszek R, Cowling VH. Oncogenic PIK3CA mutations increase dependency on the mRNA cap methyltransferase, RNMT, in breast cancer cells. Open Biol. 2019;9:190052
179. Butterfield LH, Najjar YG. Immunotherapy combination approaches: mechanisms, biomarkers and clinical observations. Nat Rev Immunol. 2023
180. Kroemer G, Chan TA, Eggermont A, Galluzzi L. Immunosurveillance in clinical cancer management. Ca-Cancer J Clin. 2023
Author contact
![]() Corresponding authors: Correspondence to He Zhu, Yonglong Pan or Bixiang Zhang. Hepatic Surgery Center, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Hubei Province for the Clinical Medicine Research Center of Hepatic Surgery, 1095 Jiefang Avenue, 430030 Wuhan, China. E-mail: tjhzhuheedu.cn, pyl941023com, bixiangzhangcom.
Corresponding authors: Correspondence to He Zhu, Yonglong Pan or Bixiang Zhang. Hepatic Surgery Center, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Hubei Province for the Clinical Medicine Research Center of Hepatic Surgery, 1095 Jiefang Avenue, 430030 Wuhan, China. E-mail: tjhzhuheedu.cn, pyl941023com, bixiangzhangcom.